Минфин ужесточает правила подачи налоговых деклараций
Налоговая декларация будет признаваться неподанной, если ее подаст человек, не уполномоченный представлять организацию. Такая новация содержится в новом законопроекте о внесении изменений в ст. 80 и 88 ч. I Налогового кодекса (НК) РФ. Основаниями для непринятия налоговой декларации также могут оказаться недостоверные сведения об организации или ее отсутствие в ЕГРЮЛе, исключение компании или ИП из реестра. В случае выявления хотя бы одного из указанных признаков в налоговой отчетности она будет считаться неподанной. Это может усложнить работу компаний, считают юристы.
Сейчас ФНС может принять или отказать в приеме при подаче декларации не в ту налоговую; без подписи ответственного лица; без документов, удостоверяющих личность ответственного физического лица. При этом перечень оснований для отказа может отличаться для деклараций в электронном и бумажном виде, говорится в приказе ФНС от 08.07.2019 № ММВ-7-19/343.
В законе эти основания не перечислены, обращают внимание юристы. «На практике отказы в приеме деклараций по основаниям, предусмотренным в проекте закона, существовали и ранее, однако их перечень был урегулирован только на уровне ведомственных актов ФНС, – поясняет партнер налоговой практики CMS Russia Гайк Сафарян. – При этом в НК РФ прямо закреплено, что налоговый орган не вправе отказать в принятии налоговой декларации, если она предоставлена налогоплательщиком в установленной для этого форме».
«На практике отказы в приеме деклараций по основаниям, предусмотренным в проекте закона, существовали и ранее, однако их перечень был урегулирован только на уровне ведомственных актов ФНС, – поясняет партнер налоговой практики CMS Russia Гайк Сафарян. – При этом в НК РФ прямо закреплено, что налоговый орган не вправе отказать в принятии налоговой декларации, если она предоставлена налогоплательщиком в установленной для этого форме».
Уже сейчас налоговая может не принять декларацию по причинам, перечисленным в законопроекте. «Например, ИФНС может отказать в приеме декларации, если она подписана лицом, у которого отсутствуют полномочия подтверждать достоверность и полноту указанных сведений», – отмечает партнер юридической компании «Генезис» Василий Сосновский. Однако чаще всего налоговики идут навстречу налогоплательщику и принимают декларации даже с изъянами, а исправить недочеты сейчас можно без риска каких-либо санкций, указывает юрист.
Законопроект Минфина направлен на борьбу с подачей притворных деклараций. «Минфин четко хочет определить в НК шесть оснований, при наличии которых налоговая инспекция сможет отказать в приеме деклараций», – поясняет Сосновский. Хотя от новых правил могут пострадать и добросовестные налогоплательщики. «На практике иногда случаются ситуации, когда работающие организации не могут сдать декларацию по НДС из-за необоснованного отказа ФНС, – приводит пример руководитель департамента бухгалтерского учета аудиторско-консалтинговой группы «Градиент Альфа» Анна Шаталова. – Такая декларация сдается исключительно в электронном формате. Если ее не принимают, организации грозит не только штраф за несвоевременную сдачу декларации, но и приостановление операций по банковским счетам (из-за несдачи декларации в течение 10 дней). Также возникнут проблемы у контрагентов-покупателей, которые не смогут подтвердить вычеты по НДС, пока не будет принята электронная декларация у организации-поставщика». При новых требованиях риск возникновения таких ситуаций может возрасти.
«Минфин четко хочет определить в НК шесть оснований, при наличии которых налоговая инспекция сможет отказать в приеме деклараций», – поясняет Сосновский. Хотя от новых правил могут пострадать и добросовестные налогоплательщики. «На практике иногда случаются ситуации, когда работающие организации не могут сдать декларацию по НДС из-за необоснованного отказа ФНС, – приводит пример руководитель департамента бухгалтерского учета аудиторско-консалтинговой группы «Градиент Альфа» Анна Шаталова. – Такая декларация сдается исключительно в электронном формате. Если ее не принимают, организации грозит не только штраф за несвоевременную сдачу декларации, но и приостановление операций по банковским счетам (из-за несдачи декларации в течение 10 дней). Также возникнут проблемы у контрагентов-покупателей, которые не смогут подтвердить вычеты по НДС, пока не будет принята электронная декларация у организации-поставщика». При новых требованиях риск возникновения таких ситуаций может возрасти.
При этом юристы обращают внимание, что предложенный перечень оснований лишь усложнит деятельность добросовестных организаций и почти не повлияет на мошеннические. «Предложенный перечень оснований недостаточный для того, чтобы выявлять новых недобросовестных налогоплательщиков, – замечает Сосновский. – Владельцы фирм-однодневок смогут его обойти, например, с помощью использования электронной цифровой подписи, оформленной на номинальных директоров».
«Предложенный перечень оснований недостаточный для того, чтобы выявлять новых недобросовестных налогоплательщиков, – замечает Сосновский. – Владельцы фирм-однодневок смогут его обойти, например, с помощью использования электронной цифровой подписи, оформленной на номинальных директоров».
Изменения в сфере налогообложения на имущество
Все конфигурации
1С:ERP Управление предприятием 2,5
1С:ERP Управление предприятием 2. 4
4
1С:Управление холдингом 8
Бухгалтерия Некоммерческой организации Корп
Бухгалтерия некоммерческой организации Проф
Бухгалтерия предприятия КОРП, редакция 2. 0
0
Бухгалтерия предприятия КОРП, редакция 3.0
Бухгалтерия предприятия, редакция 2.0
Бухгалтерия предприятия, редакция 3.0
Зарплата и управление персоналом КОРП, редакция 2. 5
5
Зарплата и управление персоналом, редакция 3
Зарплата и управление персоналом, редакция 3 LTS
Комплексная автоматизация, редакция 2,4
Комплексная автоматизация, редакция 2. 5
5
Платежные документы
Розница, редакция 2.3
Садовод
Управление нашей фирмой
Управление производственным предприятием, редакция 1. 3
3
Управление торговлей, редакция 10.3
Официальное опубликование правовых актов ∙ Официальный интернет-портал правовой информации
1.
Приказ Федеральной налоговой службы от 23. 03.2021 № ЕД-7-21/215@
03.2021 № ЕД-7-21/215@«Об утверждении формы, порядка ее заполнения, формата и порядка представления в налоговый орган по субъекту Российской Федерации сведений о кадастровых номерах земельных участков, предоставленных на праве постоянного (бессрочного) пользования федеральным органам исполнительной власти, федеральным государственным органам, в которых федеральными законами предусмотрена военная служба (приравненная к ней служба), федеральному органу исполнительной власти, уполномоченному по контролю и надзору в области таможенного дела, и находящимся в их ведении органам и иным лицам и относящихся к земельным участкам, изъятым из оборота в соответствии с законодательством Российской Федерации, и к земельным участкам, ограниченным в обороте в соответствии с законодательством Российской Федерации, предоставленным для обеспечения обороны, безопасности и таможенных нужд»
(Зарегистрирован 21.05.2021 № 63565)
PDF-файл: 2136 Кб (18 стр.
 )
Номер опубликования:
0001202105240026
Дата опубликования:
)
Номер опубликования:
0001202105240026
Дата опубликования:
Номер опубликования:
0001202105240026
Дата опубликования:
24.05.2021
2.
Номер опубликования:
0001202105200015
Дата опубликования:
20.05.2021
3.
Номер опубликования:
0001202105190031
Дата опубликования:
19.
4.
Приказ Федеральной налоговой службы от 19.02.2021 № ЕД-7-21/152@«Об утверждении формы и формата представления сведений о лицах, в отношении которых приняты решения о назначении пенсии, прекращении выплаты пенсии, о лицах, соответствующих условиям, необходимым для назначения пенсии в соответствии с законодательством Российской Федерации, действовавшим на 31 декабря 2018 года, о лицах, относящихся к ветеранам боевых действий, сведения о которых размещены в Единой государственной информационной системе социального обеспечения, а также о лицах, сведения о которых внесены в федеральный реестр инвалидов, порядка заполнения указанной формы и признании утратившим силу приказа Федеральной налоговой службы от 12.11.2019 № ММВ-7-21/567@»
PDF-файл: 2755 Кб (18 стр.
 )
Номер опубликования:
0001202105170071
Дата опубликования:
17.05.2021
)
Номер опубликования:
0001202105170071
Дата опубликования:
17.05.2021
Номер опубликования:
0001202105170071
Дата опубликования:
17.05.2021
5.
Номер опубликования:
0001202105140036
Дата опубликования:
14.05.2021
6.
Номер опубликования:
0001202104280034
Дата опубликования:
28. 04.2021
04.2021
7.
Номер опубликования:
0001202104260012
Дата опубликования:
26.04.2021
8.
Номер опубликования:
0001202104260002
Дата опубликования:
26.04.2021
9.
Номер опубликования:
0001202104230001
Дата опубликования:
23. 04.2021
04.2021
10.
Номер опубликования:
0001202104230010
Дата опубликования:
23.04.2021
11.
Номер опубликования:
0001202104210001
Дата опубликования:
21.04.2021
12.
Номер опубликования:
0001202104200065
Дата опубликования:
20. 04.2021
04.2021
13.
Номер опубликования:
0001202104120033
Дата опубликования:
12.04.2021
14.
Номер опубликования:
0001202104120046
Дата опубликования:
12.04.2021
15.
Номер опубликования:
0001202104060048
Дата опубликования:
06. 04.2021
04.2021
16.
Номер опубликования:
0001202103260018
Дата опубликования:
26.03.2021
17.
Номер опубликования:
0001202103180031
Дата опубликования:
18.03.2021
18.
Номер опубликования:
0001202103170012
Дата опубликования:
17. 03.2021
03.2021
19.
Номер опубликования:
0001202103170043
Дата опубликования:
17.03.2021
20.
Номер опубликования:
0001202103160041
Дата опубликования:
16.03.2021
21.
Номер опубликования:
0001202103120043
Дата опубликования:
12. 03.2021
03.2021
22.
Номер опубликования:
0001202103110030
Дата опубликования:
11.03.2021
23.
Номер опубликования:
0001202103020026
Дата опубликования:
02.03.2021
24.
Номер опубликования:
0001202102120022
Дата опубликования:
12. 02.2021
02.2021
25.
Приказ Федеральной налоговой службы от 14.01.2021 № ЕД-7-21/9@«Об утверждении форм документа о выявлении оснований, препятствующих применению порядка представления налоговой декларации по налогу на имущество организаций, предусмотренного пунктом 1.1 статьи 386 Налогового кодекса Российской Федерации, документа о выявлении оснований, препятствующих применению налогового вычета в соответствии с уведомлением о выбранном земельном участке, в отношении которого применяется налоговый вычет по земельному налогу, документа о выявлении оснований, препятствующих предоставлению налоговой льготы в соответствии с уведомлением о выбранном объекте налогообложения, в отношении которого предоставляется налоговая льгота по налогу на имущество физических лиц, уведомления о прекращении исчисления налога на имущество физических лиц в связи с гибелью или уничтожением объекта налогообложения, сообщения об отсутствии основания для прекращения исчисления налога на имущество физических лиц в связи с гибелью или уничтожением объекта налогообложения»
(Зарегистрирован 11.
 02.2021 № 62464)
02.2021 № 62464) PDF-файл: 392 Кб (7 стр.) Номер опубликования: 0001202102120002 Дата опубликования: 12.02.2021
Номер опубликования:
0001202102120002
Дата опубликования:
12.02.2021
26.
Номер опубликования:
0001202102040020
Дата опубликования:
04.02.2021
27.
Номер опубликования:
0001202102030037
Дата опубликования:
03.02.2021
28.
Номер опубликования:
0001202101290020
Дата опубликования:
29.01.2021
29.
Номер опубликования:
0001202101290015
Дата опубликования:
29. 01.2021
01.2021
30.
Номер опубликования:
0001202101200011
Дата опубликования:
20.01.2021
ФНС изменит форму налоговой декларации по УСН
ФНС подготовила поправки в форму налоговой декларации по УСН. Проект соответствующего приказа опубликован на Едином портале для размещения проектов НПА.
Напомним, действующая форма декларации по УСН утверждена приказом от 26.02.2016 № ММВ-7-3/99@.
Теперь ФНС предлагает изменить в декларации:
- раздел 2.1.1 «Расчет налога, уплачиваемого в связи с применением упрощенной системы налогообложения, (объект налогообложения – доходы)»;
- раздел 2.
 2 «Расчет налога, уплачиваемого в связи с применением упрощенной системы налогообложения, и минимального налога (объект налогообложения — доходы, уменьшенные на величину расходов)».
2 «Расчет налога, уплачиваемого в связи с применением упрощенной системы налогообложения, и минимального налога (объект налогообложения — доходы, уменьшенные на величину расходов)».
 2 «Расчет налога, уплачиваемого в связи с применением упрощенной системы налогообложения, и минимального налога (объект налогообложения — доходы, уменьшенные на величину расходов)».
2 «Расчет налога, уплачиваемого в связи с применением упрощенной системы налогообложения, и минимального налога (объект налогообложения — доходы, уменьшенные на величину расходов)».В указанных разделах появится новая строка «Код налоговой льготы». Она должна заполняться, если за налоговый период налогоплательщиком применялась пониженная или нулевая ставка налога. Данный показатель будет составным:
- в первой части показателя надо будет указать код налоговой льготы;
- во второй части показателя будет указываться номер или буквенное обозначение соответствующей структурной единицы (статья, часть, пункт, подпункт, абзац, иное) закона субъекта РФ, которым установлена налоговая льгота.
Порядок заполнения декларации по УСН дополняется новыми кодами налоговых льгот, устанавливаемых регионами. В данном случае речь идет о пониженных ставках налога по УСН, которые могут быть установлены законами субъектов РФ в соответствии со статьей 346. 20 НК РФ.
20 НК РФ.
ФНС рассчитывает, что обновленная форма декларации по УСН будет введена с 1 января 2021 года.
В «1С:Предприятии 8» изменения будут поддержаны после утверждения соответствующего приказа ФНС с выходом очередных версий. О сроках см. в «Мониторинге законодательства».
БУХПРОСВЕТ
По общему правилу организации и индивидуальные предприниматели, применяющие УСН, обязаны сдавать налоговые декларации по итогам каждого года (ст. 346.23 НК РФ). Организации сдают декларацию не позднее 31 марта года, следующего за истекшим налоговым периодом. ИП обязаны сдавать декларацию по УСН не позднее 30 апреля года, следующего за истекшим годом.
При прекращении предпринимательской деятельности декларация сдается не позднее 25-го числа месяца, следующего за месяцем, в котором была прекращена деятельность. Если плательщик утрачивает право на применение УСН по причине, например, превышения предельно допустимого дохода, он представляет налоговую декларацию не позднее 25-го числа месяца, следующего за кварталом, в котором было утрачено право на УСН.
Между тем, в связи с распространением в стране коронавирусной инфекции и объявлением режима нерабочего времени, общие сроки представления деклараций по УСН были перенесены на более поздние сроки. Так, в соответствии с постановлением Правительства РФ от 02.04.2020 № 409 срок сдачи декларации по УСН был перенесен на 3 месяца. Организации обязаны ее сдать до 30 июня, а ИП – до 30 июля 2020 года.
ФНС сокращает сроки проведения камеральных проверок деклараций по НДС
ФНС сокращает сроки проведения камеральных проверок деклараций по НДС
ФНС запустила новый пилотный проект по сокращению сроков камеральной проверки деклараций по НДС до одного месяца. Ранее срок проверки был 2 месяца с момента представления декларации. Особенности проведения проверки в сокращенный срок приведены в письме ФНС от 06.10.2020 № ЕД-20-15/129@.
В письме отмечается, что в отношении деклараций по НДС, в которых заявлено право на возмещение сумм налога из бюджета, предусмотренное статьей 176 НК РФ, камеральная проверка может быть завершена по истечении месяца со дня представления декларации.
Днем представления декларации по НДС назначено 25-е число месяца, следующего за истекшим налоговым периодом, либо день представления уточненной декларации по НДС.
Не позднее 10 календарных дней со дня представления декларации по НДС, налоговые органы должны осуществить оценку на соответствие следующим условиям:
представление заявления о применении заявительного порядка возмещения НДС;
сумма уплаченных налогов за три года, предшествующих налоговому периоду, за который представлена налоговая декларация по НДС, превышает сумму налога заявленной к возмещению из бюджета по такой декларации.
По истечении месяца со дня представления декларации, налогоплательщики, соответствующие указанным условиям оцениваются на одновременное соответствие следующим условиям:
отсутствие ошибок в декларации и противоречий между сведениями, содержащимися в представленных документах, либо несоответствия сведений, представленных налогоплательщиком, сведениям, содержащимся в документах, имеющихся у налогового органа, и полученным им в ходе налогового контроля, приводящих к изменению налоговых обязательств;
отсутствие противоречий между сведениями об операциях, содержащихся в декларациях по НДС, представленных налогоплательщиком и его контрагентами;
отсутствие признаков нарушений законодательства о налогах и сборах, приводящих к завышению суммы налога, заявленной к возмещению из бюджета либо к занижению суммы налога, подлежащей уплате в бюджет.

При проведении камеральной проверки оценка целесообразности проведения мероприятий налогового контроля в отношении контрагентов низкого и среднего уровня налогового риска, по которым налогоплательщиком заявлены налоговые вычеты по НДС, определяется с учетом сведений, имеющихся в распоряжении налоговой.
В ФНС уверены, что сокращение срока камеральных проверок по НДС позволит бизнесу эффективнее распоряжаться оборотными капиталами и улучшит деловой климат.
В ФНС рассказали о новых правилах оформления налогового вычета
https://ria.ru/20210423/vychet-1729562382.html
В ФНС рассказали о новых правилах оформления налогового вычета
В ФНС рассказали о новых правилах оформления налогового вычета — РИА Новости, 14.05.2021
В ФНС рассказали о новых правилах оформления налогового вычета
С 21 мая россияне смогут в упрощенном порядке получать налоговый вычет из налога на доходы физлиц (НДФЛ) по расходам на покупку жилья, земли и по индивидуальным РИА Новости, 14. 05.2021
05.2021
2021-04-23T01:31
2021-04-23T01:31
2021-05-14T21:08
ндфл
россия
налоги
жилье
федеральная налоговая служба (фнс россии)
экономика
/html/head/meta[@name=’og:title’]/@content
/html/head/meta[@name=’og:description’]/@content
https://cdn21.img.ria.ru/images/07e4/0c/18/1590691426_0:279:3072:2007_1920x0_80_0_0_49e130340831aaa78ce03627eb5ee7a7.jpg
МОСКВА, 23 апр — РИА Новости. С 21 мая россияне смогут в упрощенном порядке получать налоговый вычет из налога на доходы физлиц (НДФЛ) по расходам на покупку жилья, земли и по индивидуальным инвестиционным счетам (ИИС), рассказали «Российской газете» в Федеральной налоговой службе (ФНС).Подчеркивается, что вычет можно будет оформить автоматически, не собирая никаких документов и не посещая налоговую инспекцию. Для этого понадобится заполнить заявление в личном кабинете налогоплательщика на сайте ФНС.В налоговой службе пояснили, что на возможность получения вычета в упрощенном порядке укажет автоматически предзаполненное заявление, сформированное по итогам двадцатидневной проверки полученных от банка сведений. Налогоплательщики получат специальное сообщение об этом в личном кабинете. До появления соответствующего заявления никаких действий не требуется.Чтобы уточнить, можно ли получить налоговый вычет в упрощенном порядке, гражданам советуют обратиться в банк, с которым заключен договор на ведение ИИС или на приобретение имущества.Перечень банков-участников будет актуализироваться в специальном разделе на сайте ФНС, заключили в ведомстве.
Налогоплательщики получат специальное сообщение об этом в личном кабинете. До появления соответствующего заявления никаких действий не требуется.Чтобы уточнить, можно ли получить налоговый вычет в упрощенном порядке, гражданам советуют обратиться в банк, с которым заключен договор на ведение ИИС или на приобретение имущества.Перечень банков-участников будет актуализироваться в специальном разделе на сайте ФНС, заключили в ведомстве.
https://rsport.ria.ru/20210405/sport-1604324992.html
россия
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
2021
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Новости
ru-RU
https://ria.ru/docs/about/copyright.html
https://xn--c1acbl2abdlkab1og.xn--p1ai/
РИА Новости
internet-group@rian. ru
ru
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
https://cdn23.img.ria.ru/images/07e4/0c/18/1590691426_0:0:2732:2048_1920x0_80_0_0_370e079a86192cece0409502f976cc38.jpgРИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
ндфл, россия, налоги, жилье, федеральная налоговая служба (фнс россии), экономика
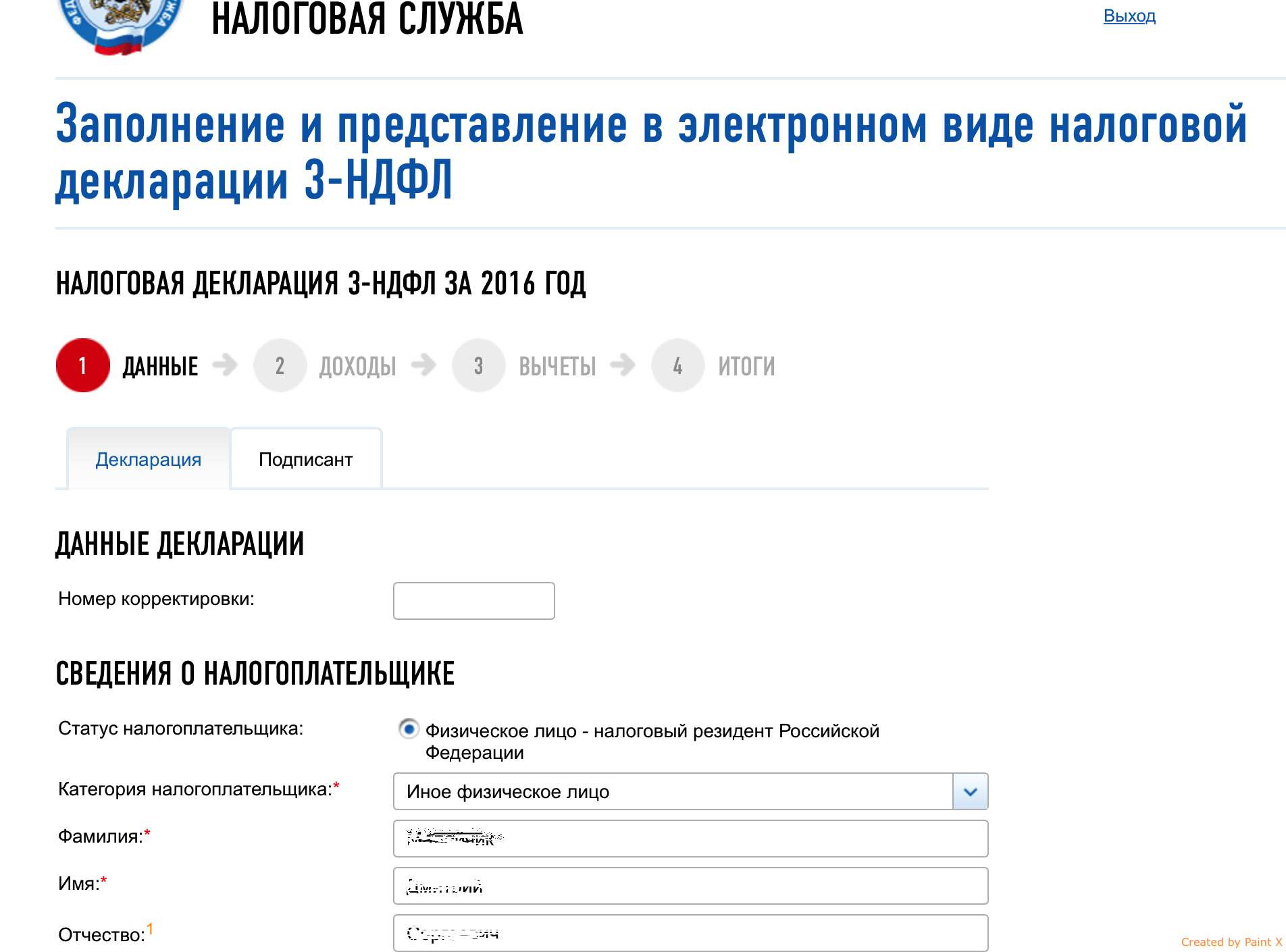
Техподдержка: Инструкция по использованию электронной подписи на сайте Федеральной налоговой службы
Оглавление
Внимание! В соответствии с пунктом 3 статьи 80 и с пунктом 5 статьи 174 Налогового кодекса Российской Федерации налоговые декларации по налогу на добавленную стоимость через сервис ФНС не принимаются. Также, согласно пункту 10 статьи 431 Налогового кодекса Российской Федерации расчеты по страховым взносам через настоящий сервис не принимаются.
Также, согласно пункту 10 статьи 431 Налогового кодекса Российской Федерации расчеты по страховым взносам через настоящий сервис не принимаются.
Установка Программы «Налогоплательщик ЮЛ»
На сайте ФНС https://www.nalog.ru/rn77/program/5961229/ скачать файл с последней версией программы и запустить установку двойным кликом мыши:
После распаковки и подготовки файлов к установке откроется окно установки программы, в котором необходимо нажать «Далее»:
Прочитать лицензионное соглашение и принять его условия, нажать «Далее»:
Выбрать «Полную» установку и нажать «Далее»:
Если необходимо, можно изменить папку установки программы, нажав «Изменить…».
Далее нажать «Далее»:
Для начала установки нажать «Установить»:
Для завершения установки программы нажать «Готово»:
После успешной установки на рабочем столе появится ярлык «Налогоплательщик ЮЛ». Также в системном меню WINDOWS Пуск – Программы появится подпункт «Налогоплательщик ЮЛ», содержащий ссылки на исполняемую программу и руководство пользователя.
При первом запуске программы после установки версии будет показано окно с описанием версии, далее выполнится конвертация программы, переиндексация, после чего появится запрос на прием отчетных форм. Затем произойдет прием описаний в выбранном варианте. После этого программа будет готова к работе. Работа в программе Налогоплательщик описана в файле «Руководство пользователя.doc» Пуск – Программы – Налогоплательщик ЮЛ – Руководство пользователя.
Формирование транспортного контейнера в программе «Налогоплательщик ЮЛ»
После формирования декларации или загрузки существующей, необходимо ее выгрузить для формирования транспортного контейнера. Для выгрузки документа нужно правым кликом мыши кликнуть на отчет. При этом если помечен документ или группа документов, то будут выгружены помеченные документы. Если нет помеченных документов, то будет выгружен документ, на котором установлен курсор.
В выпадающем меню выбрать «Передача по Интернет».
Откроется список отмеченных для выгрузки документов:
Нажать «ОК».
Откроется окно со служебной информацией:
В нем необходимо заполнить код ИФНС, в которую отправляется отчетность. И подтвердить введенные данные нажатием кнопки «ОК».
Если заполнены все необходимые поля, отчетность будет выгружена.
После нажатия кнопки «ОК» откроется окно формирования транспортного контейнера:
В нем необходимо указать папку, в которой будет размещен файл с транспортным контейнером, идентификатор налогоплательщика, сертификат ключа подписи, которым будет подписан передаваемый файл отчетности и нажать кнопку «Сформировать».
Для подписания отчетности будет запрошен пароль на контейнер:
После ввода пароля и нажатия кнопки «ОК» будет сформирован контейнер:
Получение идентификатора налогоплательщика
Для самостоятельной регистрации налогоплательщиков в системе сдачи налоговой отчетности и получения идентификатора необходимо пройти регистрацию в сервисе: https://service.nalog.ru/reg/Account/Registry:
Для регистрации необходимо ввести Логин, Пароль, Подтверждение пароля и E-mail:
После нажатия кнопки «Зарегистрировать» на электронную почту будет выслано письмо с ссылкой для подтверждения регистрации:
После подтверждения электронной почты и входа в личный кабинет появится сообщение о необходимости зарегистрировать сертификат и получить идентификатор:
После нажатия на кнопку «Зарегистрировать сертификат» откроется форма для загрузки сертификата:
После выбора файла сертификата откроется окно со сведениями об организации, в котором необходимо будет заполнить пустые поля (КПП и Код налогового органа):
Затем нажать кнопку «Передать на регистрацию». Страница обновится и отобразится статус заявки на регистрацию сертификата:
Когда сертификат пройдет регистрацию и будет присвоен идентификатор, на почту (указанную при регистрации) придет сообщение об успешной регистрации и о присвоении идентификатора.
После обновления страницы изменится статус регистрации сертификата, и в данных организации отобразится присвоенный идентификатор:
Для представления отчетности необходимо использовать «Сервис сдачи налоговой и бухгалтерской отчетности».
Представление отчетности в ФНС
Внимание! В соответствии с пунктом 3 статьи 80 и с пунктом 5 статьи 174 Налогового кодекса Российской Федерации налоговые декларации по налогу на добавленную стоимость через сервис ФНС не принимаются. Также, согласно пункту 10 статьи 431 Налогового кодекса Российской Федерации расчеты по страховым взносам через настоящий сервис не принимаются.
Для представления налоговой и бухгалтерской отчетности в электронном виде нужно зайти на страницу: http://nalog.ru/rn77/service/pred_elv/:
Далее необходимо установить Сертификат открытого ключа подписи МИ ФНС России по ЦОД, корневой сертификат ФНС России и список отозванных сертификатов.
Установка открытого ключа ФНС
Для установки сертификата открытого ключа подписи МИ ФНС России по ЦОД нужно его сохранить и запустить установку двойным кликом мыши.
На вкладке «Общие» нажать кнопку «Установить сертификат…»:
Откроется «Мастер импорта сертификатов»:
После нажатия кнопки «Далее» откроется окно выбора хранилища сертификатов.
Необходимо отметить «Автоматически выбрать хранилище на основе типа сертификата», нажать «Далее»:
Для завершения работы «Мастера импорта сертификатов» нажать кнопку «Готово»:
В окне сообщения об успешном импорте сертификата нажать кнопку «ОК»:
Сертификат открытого ключа подписи МИ ФНС России по ЦОД установлен.
Установка корневого сертификата
Для установки корневого сертификата ФНС необходимо перейти по ссылке: http://www.nalog.ru/rn77/about_fts/uc_fns/, скачать корневой сертификат УЦ ФНС России и двойным кликом мыши открыть его, для этого в окне открытия файла нажать кнопку «Открыть»:
На вкладке «Общие» нажать кнопку «Установить сертификат…»:
Откроется «Мастер импорта сертификатов»:
После нажатия кнопки «Далее» откроется окно выбора хранилища сертификатов:
Необходимо выбрать «Поместить все сертификаты в следующее хранилище», нажать кнопку «Обзор» и выбрать хранилище «Доверенные корневые центры сертификации» и нажать «ОК»:
После выбора хранилища сертификатов нажать «Далее»:
Для завершения работы «Мастера импорта сертификатов» нажать кнопку «Готово»:
В окне сообщения об успешном импорте сертификата нажать кнопку «ОК»:
Корневой сертификат установлен.
Установка списка отозванных сертификатов
Для установки списка отзыва нужно сохранить его на компьютер, кликнуть по нему правой кнопкой мыши и выбрать «Установить список отзыва (CRL)». В открывающихся окнах последовательно нажимать «Далее» – «Далее» – «Готово», не меняя настройки по умолчанию.
После установки сертификатов и списка отзыва нажать «Перейти в «Сервис сдачи налоговой и бухгалтерской отчетности».
Ознакомиться с технологией приема и обработки деклараций (расчетов) и перейти к проверке условий, нажав «Проверить выполнение условий»:
Убедиться, что все условия выполнены, и нажать «Выполнить проверки»:
На четвертом шаге проверки будет предложение выбрать цифровой сертификат.
После выбора необходимого сертификата нажать «ОК»:
После проверки сертификата ключа подписи нажать «Начать работу с сервисом»:
В открывшемся окне:
Нужно заполнить пустые поля (Код абонента, КПП) и нажать «Сохранить»:
После сохранения введенных данных перейти в раздел «Загрузка файла»:
Нажать «Обзор» и выбрать контейнер, подготовленный с помощью программы «Налогоплательщик ЮЛ».
После выбора файла нажать кнопку «Отправить».
После передачи файла произойдет автоматический переход на страницу проверки статуса обработки:
После завершения документооборота состояние изменится на «Завершено»:
Посмотреть отправленный файл и историю документооборота можно, перейдя по ссылке в графе «Состояние» – «Завершено (успешно)»:
В «Истории документооборота» можно посмотреть или скачать все регламентные документы.
В дальнейшем можно в любое время зайти в данный сервис (https://service.nalog.ru/nbo/) и просмотреть отправленные ранее декларации (расчеты).
Нарушение регуляции ответа интерферона I типа при COVID-19
Инфекция SARS-CoV-2 может привести к чрезмерной выработке провоспалительных цитокинов, но, как сообщается, производство интерферонов типа I, которые являются ключевыми противовирусными медиаторами, снижено. Здесь мы обсуждаем, как несбалансированные ответы интерферона могут способствовать патологии COVID-19.
COVID-19 характеризуется респираторным заболеванием от легкой до тяжелой степени, которое, по-видимому, зависит от возраста и сопутствующих заболеваний.У тяжелобольных пациентов часто развивается острый респираторный дистресс-синдром (ОРДС) или полиорганные поражения в результате вторичного гемофагоцитарного лимфогистиоцитоза (sHLH). И ARDS, и sHLH характеризуются чрезмерным продуцированием цитокинов и чрезмерным воспалением, что является признаком синдрома высвобождения цитокинов (CRS). CRS, вызванный SARS-CoV-2, напоминает иммунную дисрегуляцию, вызванную другими высокопатогенными респираторными вирусами, включая родственные бета-коронавирусы SARS-CoV и MERS-CoV.Заметно повышенные уровни провоспалительных цитокинов в плазме, включая IL-6 и фактор некроза опухоли (TNF), а также некоторых хемокинов были обнаружены у пациентов с COVID-19 (ref. 1 ).
Инфекции SARS-CoV-2, передающиеся воздушно-капельным путем, у людей возникают в результате проникновения вируса в эпителиальные клетки носовых и дыхательных путей через связывание с ангиотензин-превращающим ферментом 2 (ACE2). TMPRSS2, клеточная протеаза, которая активирует спайковый белок SARS-CoV-2, колокализуется с ACE2 и может инициировать слияние SARS-CoV-2 непосредственно на плазматической мембране.В легких SARS-CoV-2 поражает альвеолярные эпителиальные клетки I и II типа, а также альвеолярные макрофаги, которые являются одними из первых продуцентов провоспалительных цитокинов. В качестве ключевых компонентов немедленного противовирусного ответа интерфероны типа I (далее именуемые IFN) имеют решающее значение для ограничения репликации и распространения вируса посредством аутокринной и паракринной передачи сигналов рецептора IFN типа I (IFNAR). Однако минимальные количества IFN были обнаружены в периферической крови или легких пациентов с тяжелой формой COVID-19 (ссылки 1,2 ), что контрастирует с тем, что наблюдается у пациентов, инфицированных высокопатогенными вирусами гриппа.Интересно, что хотя низкие уровни системной продукции IFN, по-видимому, коррелируют с тяжелым COVID-19 (ссылка 2 ), локальная индукция IFN и IFN-стимулированных генов (ISG) была заметна в бронхоальвеолярном лаваже (БАЛ) некоторых тяжелобольные 3 . Это было связано с активацией специализированных иммунных клеток, таких как резидентные дендритные клетки (ДК) в легких. В частности, было показано, что плазмацитоидные DC продуцируют IFNα в ответ на SARS-CoV. У пациентов с SARS, которые не получали кортикостероиды, IFNα был обнаружен в плазме во время «предкризисной» фазы, но снизился во время «кризисной» фазы 4 .В мышиной модели инфекции SARS-CoV локальные ответы IFN в легких были задержаны относительно пика вирусной репликации, что препятствовало выведению вируса и было связано с развитием CRS 5 . Кинетика системного и местного ответа на IFN, которые возникают во время COVID-19, еще предстоит полностью выяснить, а также их соответствующий вклад в патогенез COVID-19 и тяжесть заболевания.
Нарушение регуляции ответов IFN свидетельствует об эффективных иммуномодулирующих стратегиях, используемых бета-коронавирусами.Во время фазы инкубации SARS-CoV-2 незаметно реплицируется в клетках-хозяевах, не вызывая детектируемого запуска IFN, что приводит к высокой вирусной нагрузке 1 . Известно, что коронавирусы индуцируют образование мембранных компартментов, предназначенных для синтеза вирусной РНК, и тем самым скрывают молекулярные паттерны, связанные с вирусными патогенами (PAMP; например, вирусные РНК), от обнаружения рецепторами распознавания паттернов хозяина (PRR), такими как RIG-I и MDA5. Кроме того, известно, что некоторые консервативные белки бета-коронавируса, преимущественно неструктурные белки (nsps), обладают прямой антагонистической активностью в отношении IFN.Некоторые модифицируют специфические особенности вирусной РНК (катализируя метилирование гуанозина-N7 и рибозы-2ʹ- O ), чтобы избежать распознавания специфическими PRR (например, nsp14 и nsp16), в то время как другие, такие как nsp3 и nsp1, ингибируют передача сигнала, опосредованная PRR и IFNAR, соответственно 5 . Напротив, было показано, что нуклеокапсидный белок SARS-CoV напрямую активирует NF-κB. Устойчивое производство провоспалительных цитокинов и хемокинов с ограниченным образованием IFN во время инфекции SARS-CoV-2 предполагает эффективную активацию NF-κB, но не активацию IFN-регуляторного фактора 3 (IRF3) и IRF7 (см. 1 ). Будет важно точно определить, как SARS-CoV-2 противодействует индукции IFN и передаче сигналов IFNAR.
В качестве центрального связующего звена между врожденной и адаптивной иммунной системами IFNs необходимы для регулирования активации и функций различных популяций иммунных клеток. Важно отметить, что во время инфекции SARS-CoV или MERS-CoV у мышей IFN напрямую регулируют легочную инфильтрацию макрофагов, происходящих из моноцитов. В то время как блокирование передачи сигналов IFNAR заметно снижает инфильтрацию макрофагов, отсроченная индукция IFN с помощью SARS-CoV приводит к накоплению высокоактивированных макрофагов в легких, что вызывает иммунопатологию 5 .Напротив, ингибирование IFNAR усиливало рекрутирование нейтрофилов в легкие у мышей, инфицированных MERS-CoV, что приводило к повышенной продукции провоспалительных цитокинов 6 . Нарушение выработки IFN во время тяжелой формы COVID-19 также может привести к дисбалансу провоспалительных и про-репаративных функций макрофагов дыхательных путей. У пациентов, умерших от SARS-CoV, наблюдалось накопление провоспалительных макрофагов, но дефицит макрофагов для заживления ран в легких; это было связано с более высокими уровнями нейтрализующих антител в сыворотке против спайкового белка SARS-CoV 7 .Другие клетки врожденного иммунитета, такие как естественные клетки-киллеры (NK), также регулируются IFN во время коронавирусной инфекции. Ингибирование передачи сигналов IFNAR подавляло накопление NK-клеток в легких мышей 6 , инфицированных MERS-CoV, что может замедлить ранний клиренс инфицированных вирусом клеток. В то время как пациенты с тяжелой формой COVID-19 демонстрировали глубокое истощение и функциональное истощение NK-клеток 8 , неясно, связана ли эта дисфункция NK-клеток с нарушением регуляции ответов IFN.
Тяжелая форма COVID-19 связана с нарушением Т-клеточного ответа, которое проявляется в лимфопении и функциональном истощении CD4 + и CD8 + Т-клеток 8 . Нарушение ответа Т-клеток может быть результатом недостаточной продукции ИФН, поскольку ИФН способствуют выживанию и эффекторным функциям Т-клеток. Блокирование передачи сигналов IFNAR во время инфекции БВРС-КоВ ослабляло развитие вирус-специфичных CD4 + и CD8 + Т-клеток у мышей 6 .Хотя ранняя продукция IFN имеет решающее значение для эффективного ответа Т-клеток, отсроченный ответ IFN может подавлять пролиферацию Т-клеток или выход Т-клеток из лимфоидных органов или может вызывать функциональное истощение и гибель Т-клеток. Поражение легких, связанное с СВК, у пациентов с тяжелой формой COVID-19 указывает на возможную неспособность своевременно активировать иммуносупрессивные механизмы. Действительно, было показано, что количество регуляторных Т-клеток (T reg ) у пациентов с COVID-19 обратно коррелирует с тяжестью заболевания 9 .Известно, что IFN являются ключевыми регуляторами развития клеток T reg . Таким образом, возникает соблазн предположить, что недостаточный или нерегулируемый ответ IFN, вызванный инфекцией SARS-CoV-2, может влиять на генерацию клеток T reg во время фазы восстановления COVID-19. В будущих исследованиях следует изучить, как нарушение регуляции IFN во время COVID-19 может формировать Т-клеточные ответы, и, учитывая, что активация Т-клеток CD4 + имеет решающее значение для развития В-клеточного иммунитета, как это, в свою очередь, может повлиять на ответы антител.
То, что нарушение регуляции IFN представляет собой ключевой детерминант патогенеза COVID-19, подчеркивает его потенциал для терапевтического вмешательства. Профилактическое введение IFN, которое вызывает ранее существовавшее противовирусное состояние в клетках-мишенях, может блокировать вирусную инфекцию на самой ранней стадии. Было показано, что ежедневные назальные капли с IFNα вместе со стандартными средствами индивидуальной защиты (СИЗ) защищают медицинских работников из группы риска от COVID-19 в течение 28 дней без заметных побочных эффектов (NCT04320238).Использование интерферонов для лечения COVID-19 остается спорным, особенно в отношении сроков приема. Раннее лечение IFN до пика вирусной репликации защищало мышей от смертельного заражения SARS-CoV или MERS-CoV, тогда как позднее введение IFN затрудняло клиренс вируса и усугубляло иммунопатологию 5,6 . Клинические исследования SARS-CoV и MERS-CoV также показали неубедительные эффекты IFN в сочетании с противовирусными препаратами на исходы заболевания, что, вероятно, связано с разным временем приема, а также сопутствующими заболеваниями.Важно отметить, что ACE2 был недавно идентифицирован как ISG в эпителиальных клетках дыхательных путей человека 10 . Может ли профилактическое или терапевтическое введение IFN усилить проникновение и репликацию SARS-CoV-2 в клетках-мишенях во время прогрессирования заболевания, является потенциальной проблемой безопасности. Дальнейшие исследования должны также определить вклад генетики хозяина, возраста и сопутствующих заболеваний в терапевтическую эффективность интерферонов. В то время как несколько текущих клинических испытаний оценивают эффективность лечения IFN для COVID-19, более глубокое понимание пространственно-временной кинетики ответов IFN во время клинических инфекций SARS-CoV-2 необходимо для получения информации о терапии, связанной с IFN, и разработке вакцины.
Ссылки
- 1.
Blanco-Melo, D. et al. Несбалансированная реакция хозяина на SARS-CoV-2 способствует развитию COVID-19. Ячейка https://doi.org/10.1016/j.cell.2020.04.026 (2020).
Артикул PubMed PubMed Central Google Scholar
- 2.
Hadjadj, J. et al. Нарушение активности интерферона I типа и обострение воспалительных реакций у пациентов с тяжелым Covid-19.Препринт на medRxiv https://doi.org/10.1101/2020.04.19.20068015 (2020).
Артикул Google Scholar
- 3.
Zhou, Z. et al. Повышенный врожденный иммунный ответ в дыхательных путях у пациентов с COVID-19. Cell Host Microbe https://doi.org/10.1016/j.chom.2020.04.017 (2020).
Артикул PubMed PubMed Central Google Scholar
- 4.
Cameron, M. J. et al. Опосредованные интерфероном иммунопатологические явления связаны с атипичными врожденными и адаптивными иммунными ответами у пациентов с тяжелым острым респираторным синдромом. J. Virol. 81 , 8692–8706 (2007).
CAS Статья Google Scholar
- 5.
Channappanavar, R. et al. Нарушение регуляции интерферона I типа и воспалительные реакции моноцитов-макрофагов вызывают летальную пневмонию у мышей, инфицированных SARS-CoV. Клеточный микроб-хозяин 19 , 181–193 (2016).
CAS Статья Google Scholar
- 6.
Channappanavar, R. et al. Время ответа IFN-I относительно репликации вируса определяет исходы заражения коронавирусом MERS. J. Clin. Вкладывать деньги. 130 , 3625–3639 (2019).
Артикул Google Scholar
- 7.
Liu, L. et al.IgG к спайку вызывает тяжелое острое повреждение легких, искажая ответы макрофагов во время острой инфекции SARS-CoV. JCI Insight 4 , e123158 (2019).
Артикул Google Scholar
- 8.
Zheng, M. et al. Функциональное истощение противовирусных лимфоцитов у пациентов с COVID-19. Cell Mol. Иммунол. 17 , 533–535 (2020).
CAS Статья Google Scholar
- 9.
Qin, C. et al. Нарушение регуляции иммунного ответа у пациентов с коронавирусом 2019 (COVID-19) в Ухане, Китай. Clin. Заразить. Дис. https://doi.org/10.1093/cid/ciaa248 (2020).
Артикул PubMed PubMed Central Google Scholar
- 10.
Ziegler, C.G.K. et al. Рецептор SARS-CoV-2 ACE2 представляет собой стимулируемый интерфероном ген в эпителиальных клетках дыхательных путей человека и обнаруживается в определенных подмножествах клеток в тканях. Ячейка https://doi.org/10.1016/j.cell.2020.04.035 (2020).
Артикул PubMed PubMed Central Google Scholar
Скачать ссылки
Информация об авторе
Принадлежности
Департамент микробиологии, Чикагский университет, Чикаго, Иллинойс, США
Дхирадж Ачарья, ГуанКун Лю и Микаэла У. Гэк
000
9 Вклады2 Авторы одинаково внесли свой вклад во все аспекты статьи.
Автор, ответственный за переписку
Микаэла У. Гак.
Декларации этики
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Ссылки по теме
ClinicalTrials.gov: https://clinicaltrials.gov/
Об этой статье
Цитируйте эту статью
Acharya, D., Лю Дж. И Гак М.У. Нарушение регуляции ответа интерферона I типа при COVID-19. Nat Rev Immunol 20, 397–398 (2020). https://doi.org/10.1038/s41577-020-0346-x
Скачать цитату
Дополнительная литература
Взаимодействие между воспалительными путями и COVID-19: критический обзор патогенеза и терапевтических возможностей
- Шалки Чоудхари
- , Каджал Шарма
- и Ом Силакари
Микробный патогенез (2021 год)
Прототип QSP-модели иммунного ответа на SARS ‐ CoV ‐ 2 для развития сообщества
- Вэй Дай
- , Рохит Рао
- , Анна Шер
- , Несси Таня
- , Синтия Дж.Мусанте
- и Ричард Аллен
CPT: Фармакометрия и системная фармакология (2021 год)
Синдром острого респираторного дистресс-синдрома и COVID-19 у ребенка с системной красной волчанкой
- Симоне Сухдео
- , Эмили Негропонте
- , Харипрем Раджасекхар
- , Сунанда Гаур
- , Даниэль Б. Хортон
- , Амиша Малхотра
- и Л. Нандини Мурти
Волчанка (2021 год)
Почему COVID-19 более серьезен у пациентов с диабетом? Роль ангиотензин-превращающего фермента 2, эндотелиальной дисфункции и иммуновоспалительной системы
- Джейкоб Робертс
- , Антония Л.Причард
- , Эндрю Т. Тревик
- , Адриано Г. Росси
- , Николь Брейс
- , Пол Кэхилл
- , Сандра М. Макрури
- , Джун Вей
- и Ян Л. Мегсон
Границы сердечно-сосудистой медицины (2021 год)
Клиническое и научное обоснование больничного протокола лечения COVID-19 «MATH +»
- Пьер Кори
- , Г.Умберто Медури
- , Хосе Иглесиас
- , Джозеф Варон
- и Пол Э. Марик
Журнал интенсивной терапии (2021 год)
Интерферон-бета предлагает многообещающие возможности лечения COVID-19: систематический обзор и метаанализ клинических исследований
Путем поиска в 4 базах данных мы обнаружили 356 записей. Полученные записи были оценены на предмет дублирования, а заголовки и аннотации были проанализированы в соответствии с критериями включения.Из 356 записей 12 РКИ и когортных исследований были отобраны для полнотекстового скрининга. Наконец, 5 записей были включены в систематический обзор, а 3 статьи — в метаанализ.
Всего участников было 314 пациентов. На блок-схеме PRISMA показаны включенные исследования (рис. 1). После полнотекстового скрининга было исключено 7 записей (альфа-интерферон = 5, дизайн исследования (протокол) = 1, ретроспективное исследование = 1).
Рис. 1Базовые символы
В таблице 1 показаны детали включенных исследований.Мужчины были основными участниками всех исследований. Доля пациентов мужского пола колебалась от 54,0% до 75,0% (медиана 60,0%). Средний возраст участников в большинстве исследований был выше 55 лет.
Таблица 1 Включены клинические исследования и исходные характеристики среди пациентов с COVID-19, получавших IFN-βКлинические симптомы
Во всех статьях были исследованы госпитализированные пациенты с COVID-19 от легкой до тяжелой степени. В группе вмешательства доля госпитализаций в отделения интенсивной терапии (ОИТ) была ниже, чем в контрольных группах.В исследовании Monfared et al. из Ирана доля госпитализированных в ОИТ составила 45,23% и 58,97% в группах вмешательства и контрольной группе соответственно (Davoudi-Monfared et al. 2020). Кроме того, о таком же результате сообщили Rahmani et al. в исследовании РКИ по количеству госпитализированных в ОИТ (42,42% против 66,66%) (Rahmani et al.2020). Аналогичным образом, в РКИ, проведенном в Гонконге, Fan-Ngai Hung et al. сообщили, что время (в днях) раннего предупреждения с оценкой 0 (согласно национальной шкале раннего предупреждения 2) составило 4.0 и 8,0 дней для интервенционной и контрольной групп соответственно. Они продемонстрировали, что существует значимая взаимосвязь между вмешательством INF-β и снижением количества госпитализаций в ОИТ и нулевым показателем времени раннего предупреждения. Более того, лечение комбинацией противовирусных препаратов значительно снизило уровни IL-6, в то время как оно не показало какого-либо значительного влияния на концентрации TNFα и IL-10 (Hung et al., 2020) (Таблица 2).
Таблица 2 Исход и клинические показатели среди пациентов с COVID-19, получавших IFN-βУ всех пациентов во всех включенных исследованиях при поступлении наблюдалась лихорадка.В группе вмешательства у большинства участников лихорадка исчезла в конце периода исследования. В большинстве исследований количество лейкоцитов и лимфоцитов было увеличено в экспериментальной группе. Исследование Dastan et al. продемонстрировали, что лихорадка у всех пациентов исчезла в течение первых семи дней. Аналогичным образом, лихорадка исчезла у 81% и 78% пациентов в группе вмешательства и в контрольной группе соответственно. Более того, сравнение КТ легких и рентгенографии грудной клетки при поступлении и на 14-й день показало непрозрачность матового стекла у 16 пациентов и двустороннюю инфильтрацию у 14 пациентов, что продемонстрировало наступление выздоровления через 14 дней (Dastan et al.2020).
Время появления симптомов от начала лечения составляло от 5 до 10 дней. О самом коротком времени введения IFN после появления симптомов сообщил Гонконг. Это исследование не показало смертности в конце исследования. О самом продолжительном времени от появления симптомов до введения интерферона сообщили Monfared et al. в Иране; в конце этого исследования показатели смертности в группах вмешательства и контроля составили 19% и 43,6% соответственно (Davoudi-Monfared et al.2020).
Вмешательство (доза IFN-β)
В большинстве исследований 44 мкг / мл (12 миллионов МЕ / мл) INF-β вводили три раза в неделю в течение двух недель подряд или до выписки. Во всех исследованиях INF-β вводили подкожно. Более того, в большинстве исследований не сообщалось о серьезных побочных эффектах терапии интерфероном среди групп вмешательства. Только в многоцентровом РКИ, проведенном в Гонконге, исследователи сообщили о 2% побочных эффектах терапии интерфероном; однако они в основном были легкими и самоограничивающимися (Hung et al.2020).
Исходы
Мы обнаружили, что показатели исходов (выписка и уровень смертности) были снижены во всех исследованиях в группе вмешательства. Среднее количество дней госпитализации в обеих исследуемых группах колебалось от 6,75 до 16,8 дней; однако средняя продолжительность госпитализации в группах вмешательства (9 дней) была ниже, чем в контрольных группах (12,25 дня).
Что касается уровня смертности, Rahmani et al. сообщили, что смертность и выписки составили 6,06% против 18,18% и 78,79% против 54.55% в группах IFN по сравнению с контрольной группой, соответственно (Rahmani et al.2020). В другом исследовании (Dastan et al.) Не было контрольной группы для сравнения результатов. Аналогичным образом, в одном исследовании во время исследования в группах вмешательства и контрольной группе не было летальных исходов.
Однако средний уровень смертности в группе INF-β и стандартной группе составил 6,195% и 18,02% соответственно.
В многоцентровом РКИ Fan-Ngai Hung et al. в Гонконге среднее количество дней госпитализации в экспериментальной и контрольной группах составило 9.0 и 14,5 суток соответственно. Кроме того, в конце исследования они не сообщили о смерти в экспериментальной и контрольной группах (Hung et al., 2020). В рандомизированном контролируемом исследовании, проведенном в Иране, уровень смертности в экспериментальной и контрольной группах составил 19% и 43,6% в течение 28 дней периода исследования. Напротив, в этом исследовании сообщается, что продолжительность госпитализации в группе вмешательства была больше, чем в контрольной группе (14,80 ± 8,45 против 12,25 ± 7,48) (Davoudi-Monfared et al.2020). В другом открытом клиническом исследовании с участием одной группы сообщалось только об одной смерти после 45 дней госпитализации (Payandemehr et al.2020). В другом неконтролируемом исследовании лихорадка исчезла у всех пациентов в течение первых 7 дней (Dastan et al., 2020).
Мета-анализ
Из четырех включенных исследований три исследования имели размер эффекта (HR и OR) для вероятности выписки для сравнения терапевтических эффектов INF-β среди исследуемых групп (группа стандартной помощи и группа INF-β). Только в одном исследовании сообщалось о степени воздействия на уровень смертности. Поэтому мы провели метаанализ для оценки распространенности выписанных пациентов среди трех исследований.Во всех исследованиях частота выписки была достоверно связана с терапией INF-β. Согласно метаанализу, значительная разница была обнаружена между группами вмешательства и контрольной группой по общей скорости выпадения (ОР = 3,05; 95% ДИ, (1,09–5,01). Не было значительной гетерогенности ( P = 0,46) (рис. 2).
Рис. 2Эффекты терапии интерфероном β-1 у пациентов с COVID-19
Frontiers | Функциональное истощение выработки интерферонов типа I и II у пациентов с тяжелой формой COVID-19
Введение
В начале декабря 2019 г. первые случаи вирусной пневмонии неизвестного происхождения были выявлены в Ухане, столице провинции Хубэй в Китае (1, 2).Вирус был идентифицирован как новый бета-коронавирус, который теперь называется коронавирусом 2 тяжелого острого респираторного синдрома (SARS-CoV-2) из того же семейства, что и SARS-CoV, ответственный за вспышку атипичной пневмонии в 2003 году. Этот коронавирус, вызывающий новое коронавирусное заболевание. (COVID-19) с тех пор распространился по миру и вызвал пандемию (3, 4).
Общие симптомы у пациентов с COVID-19 включают лихорадку, сухой кашель, аносмию, одышку и другие симптомы гриппа (3–5). Несмотря на то, что у большинства пациентов могут оставаться полностью бессимптомные симптомы или могут проявляться только легкие симптомы, у 10–20% пациентов развивается тяжелое заболевание, характеризующееся тяжелой пневмонией, острым респираторным дистресс-синдромом и полиорганной недостаточностью, требующей немедленной госпитализации в отделения интенсивной терапии и часто приводит к летальному исходу (3, 6, 7).Тяжелые клинические симптомы, такие как диффузное поражение альвеол, тромбоз, гемофагоцитоз и истощение иммунных клеток, были описаны в подгруппе пациентов с тяжелой формой COVID-19 (8). Пациенты, страдающие диабетом, раком или другими хроническими заболеваниями, наиболее подвержены риску развития тяжелой формы (9).
Чтобы лучше разделить пациентов, которые могут подвергаться риску осложнений, многочисленные исследования выявили биологические маркеры с худшим прогнозом, такие как лимфопения, и воспалительные маркеры, такие как C-реактивный белок (CRP), лактатдегидрогеназа (LDH) и уровни цитокинов (3). , 10–15).
Несколько авторов исследовали роль различных цитокинов у пациентов с COVID-19. В рамках иммунологического ответа на инфекцию SARS-CoV-2 пациенты часто проявляют агрессивный и неконтролируемый воспалительный ответ с секрецией большого количества провоспалительных цитокинов, таких как интерлейкин (IL) 6, IL10 и фактор некроза опухоли α. (TNFα) в событии, известном как цитокиновый шторм (3, 16–22). Цитокиновый шторм напрямую коррелирует с повреждением легких, полиорганной недостаточностью и неблагоприятным прогнозом (19).
Взаимодействие между врожденным и адаптивным иммунным ответом, по-видимому, имеет решающее значение для определения эволюции пациента, характеризующейся дисбалансом про- и противовоспалительных цитокинов и последующим нарушением регуляции иммунного ответа пациента (23). Интерфероны (IFN) действуют как ключевое звено между врожденным и адаптивным иммунным ответом. IFN типа I (IFN-α / β) секретируются плазматическими дендритными клетками (pDC), в то время как IFN типа II (IFN-γ) преимущественно продуцируются естественными клетками-киллерами и в меньшей степени Т-клетками и макрофагами (24–26) .ИФН типа I и типа II обладают множеством противовирусных эффектов, таких как индукция апоптоза инфицированных клеток и активация макрофагов, естественных киллеров (NK) и Т-лимфоцитов (24–26). Несколько исследований показали нарушение регуляции выработки интерферонов у пациентов с COVID-19 (14, 27, 28).
В поисках новых и мощных биомаркеров неблагоприятного исхода у пациентов с COVID-19 мы проанализировали возможности ответа иммунной системы с помощью in vitro, стимуляции как клеток адаптивного, так и врожденного иммунитета, таким образом эффективно имитируя вирусную инфекцию.Действительно, ранее было показано, что стимуляция in vitro и клеток врожденного и адаптивного иммунитета позволяет прогнозировать худший исход при других заболеваниях, связанных с иммунитетом (29, 30), но, насколько нам известно, еще не исследовалась у пациентов с COVID-19. В этом проспективном одноцентровом исследовании мы сравнили функцию клеток врожденного и адаптивного иммунитета у пациентов с COVID-19 с легкой, умеренной или тяжелой формой заболевания, чтобы подчеркнуть механизм, ответственный за нарушение регуляции иммунного ответа.
Материалы и методы
Дизайн исследования и популяция
Мы провели проспективное когортное исследование в университетской больнице Ниццы. Критериями включения были: (1) все взрослые пациенты, поступившие по поводу COVID-19 в консультационное отделение (отделение дерматологии или инфекционных заболеваний), в отделения инфекционных болезней или в отделение интенсивной терапии университетской больницы Ниццы с марта по апрель 2020 года; (2) отсутствие иммуносупрессивной терапии в течение 6 месяцев до включения; (3) возможность подписать информированное согласие.Критерии исключения: (1) все пациенты до 18 лет; (2) пациенты, находящиеся под стражей, в тюрьме или страдающие психическим заболеванием; (3) беременные или кормящие грудью; (4) с известным иммунодефицитом или получавшим ранее иммуносупрессивную терапию. Было также набрано 50 здоровых доноров, не инфицированных SARS-CoV-2 (подтверждено отрицательным серологическим тестом).
В зависимости от тяжести заражения SARS-CoV-2 пациенты были разделены на три группы: (а) пациенты с тяжелой формой COVID-19 — госпитализированные или переведенные в отделение интенсивной терапии с респираторным дистресс-синдромом или дыхательной недостаточностью. требующие искусственной вентиляции легких или полиорганной недостаточности; (b) пациенты с COVID-19 средней тяжести — это пациенты, госпитализированные в отделения инфекционных болезней, в зависимости от клинических симптомов, связанных с одышкой, и радиологических данных о пневмонии при КТ грудной клетки; (c) Пациенты с COVID-19 с легкими симптомами COVID, такими как обморожение пальцев рук и ног или гриппоподобные симптомы, не требующие наблюдения в больнице.Все пациенты имели симптоматику COVID-19 в соответствии с классификацией ВОЗ с помощью компьютерной томографии, характерной для COVID-19 (31) или обморожения (32), или двух последовательных положительных результатов ОТ-ПЦР-теста на SARS-CoV-2 на образцах из верхних и нижних дыхательных путей. (мазок из носоглотки или инвазивный респираторный образец) или положительный серологический тест (Euroimmun® ELISA).
Эпидемиологические, биологические и клинические данные на день 0 (D0) представлены в таблице 1 и дополнительной таблице 1. Лечение (а), полученное после D0, обобщено в дополнительной таблице 2.Осложнения определялись как все нежелательные явления, такие как госпитализация в отделение интенсивной терапии после ухудшения симптомов, искусственная вентиляция легких, тромбоз глубоких вен, вторичная бактериальная инфекция, почечная недостаточность, гепатит, сердечная недостаточность и смерть.
Таблица 1 . Демографические и исходные характеристики здоровых доноров и пациентов с COVID-19.
Информированное согласие было получено для всех пациентов. Протокол исследования соответствовал этическим принципам Хельсинкской декларации 1975 года и был одобрен соответствующим институциональным комитетом по обзору (NCT04355351).
Сбор крови и анализ цитокинов
Образцы крови были собраны в D0 и в несколько контрольных точек в срок до 2 месяцев после поступления в больницу. Один миллилитр цельной крови стимулировали иммунными лигандами (анти-CD3 в качестве стимулятора Т-клеток и R848 в качестве агониста Toll-подобных рецепторов 7/8 (TLR 7/8)) на отдельных лиофилизированных сферах (LyoSphere TM , Qiagen) в течение 8 ч после сбора крови. Образцы стимулированной крови инкубировали в течение 16–24 ч при 37 ° C, а затем центрифугировали при 2000–3000 × g в течение 15 мин для сбора стимулированной сыворотки.Нестимулированные сыворотка и плазма, а также стимулированная сыворотка хранили при -20 ° C до минимизации циклов анализа и замораживания-оттаивания для сохранения качества образцов. Уровни цитокинов в сыворотке и плазме крови с неспецифической стимуляцией или без нее измеряли с помощью теста QuantiFERON-Monitor для обнаружения IFN-γ или специально разработанных картриджей Ella (ProteinSimple) для обнаружения IL-1β, IL-6, IL-8, IL-10, IL-17A, TNF-α и IFN-α в соответствии с инструкциями производителя.
Исследования
in vitroДля 18 пациентов с COVID-19 один миллилитр цельной крови, взятой в D0, был предварительно обработан различными молекулами в течение 6 часов при 37 ° C с последующей стимуляцией иммунными лигандами на отдельных лиофилизированных сферах (LyoSphere TM , Qiagen), как описано в разделе «Сбор крови и анализ цитокинов».Использовались молекулы, которые обычно вводили пациентам с COVID-19 (33–43): гидроксихлорохин (100 мкМ, Inresa), тоцилизумаб против IL6 (100 мкг / мл, RoActemra, Roche), метилпреднизолон (20 мкг / мл, Mylan). , адалимумаб против TNFα (10 мкг / мл, Humira, AbbVie), рекомбинантный человеческий IL-2 (6 нг / мл, Sigma), рекомбинантный человеческий IFN-альфа (100 нг / мл, Sigma) и ниволумаб (1 мкг / мл , Opdivo, Bristol Myers Squibb).
Статистика
Для описательной статистики данные представлены в виде среднего и стандартного отклонения для непрерывных значений с гауссовым распределением, в виде медианы и диапазона для непрерывных значений с негауссовым распределением, а также в виде количества и процентов для категориальных переменных.Тест нормальности Д’Агостино и Пирсона использовался для определения того, имеет ли переменная гауссово распределение или нет. Группы непрерывных значений сравнивали с помощью теста Манна-Уитни, однофакторного дисперсионного анализа ANOVA (> 2 групп) или теста Краскела-Уоллиса (> 2 групп). Множественные сравнительные тесты были выполнены с тестом Краскела-Уоллиса с использованием апостериорного теста Данна . Категориальные переменные сравнивались с использованием критерия хи-квадрат. Кривая AUC (площадь под кривой) ROC (рабочая характеристика приемника) использовалась для определения порога IFN-γ, который наилучшим образом определяет пациентов с осложнениями или без них.Для сравнения данных о выживаемости использовался лог-ранговый тест. Для сравнения двух измерений непрерывной переменной, выполненных на одних и тех же испытуемых (парные данные), использовался знаковый ранговый критерий Уилкоксона. Логистическая регрессия была проведена для определения отношений ODDS и 95% доверительных интервалов (ДИ). В многомерной модели мы скорректировали возраст, пол и ИМТ.
Статистический анализ выполняли с использованием GraphPad Prism 7.0 (GraphPad Software, Inc., Сан-Диего, Калифорния) или SAS 9.4. Все сравнения были двусторонними, и различия считались значимыми, когда значение P <0.05.
Результаты
Когорта исследования
Всего 101 пациент с симптоматикой инфекции COVID-19 (таблица 1) был включен и разделен на три группы в зависимости от тяжести их симптомов на легкие ( n = 41), умеренные ( n = 30). и тяжелые случаи ( n = 30), как описано в Методах. Также было привлечено 50 здоровых доноров. Как описано ранее (1, 3, 4), наблюдалась значительная разница в возрасте, поле, ИМТ и количестве сопутствующих заболеваний среди трех групп пациентов ( p <0.0001, p = 0,0002, p = 0,0003 и p = 0,0018 соответственно). Наиболее частые симптомы инфекции COVID-19 включали кашель, одышку и лихорадку у 52, 52 и 42% пациентов соответственно (дополнительная таблица 1).
Уровни цитокинов в нестимулированной плазме
Как и ожидалось, более высокие уровни провоспалительных цитокинов IL1β, IL6, IL8 и TNFα в плазме крови при поступлении и до специального лечения положительно коррелировали с тяжестью симптомов COVID-19 ( p <0.0001 для всех цитокинов) (таблица 2), что подтверждает результаты предыдущих исследований (3, 16–18).
Таблица 2 . Исходные уровни нестимулированных цитокинов в плазме здоровых доноров и пациентов с COVID-19.
Уровни цитокинов в сыворотке крови после
in vitro Неспецифическая стимуляция клеток врожденного и адаптивного иммунитетаХотя текущее состояние воспалительного ответа на инфекцию SARS-CoV-2, о чем свидетельствуют уровни цитокинов в плазме, отражает продолжающееся взаимодействие между врожденным и адаптивным иммунитетом, оно мало что говорит нам об иммунной функции.С этой целью мы стимулировали врожденные клетки и Т-лимфоциты пациентов с COVID-19 при поступлении и перед специфическим лечением агонистом Toll-подобного рецептора 7/8 (TLR 7/8) и анти-CD3, соответственно, и измеряли секретируемые цитокины. . TLR7 преимущественно экспрессируется в плазматических дендритных клетках (pDC) (44), а TLR8 сильнее экспрессируется в миелоидных дендритных клетках, моноцитах и в меньшей степени в pDC (45). Цитокин Th27 IL17A, а также IFN типов I и II не обнаруживались в нестимулированной плазме пациентов с COVID-19 (таблица 2).Однако после стимуляции in vitro иммунных клеток между пациентами с разной степенью тяжести COVID-19 возникли значительные различия в уровнях цитокинов (таблица 3, рисунок 1), что отражает работоспособность их иммунной системы. Что касается врожденного иммунитета, то ДК и NK-клетки умеренных и тяжелых пациентов были функционально истощены, о чем свидетельствуют более низкие уровни IFNα (и IFNγ из NK-клеток) после стимуляции in vitro ( p <0,0001) (Рисунок 1A), поскольку ранее (46), и различия оставались значимыми после поправки на количество моноцитов ( p <0.0001 легкое против умеренного и легкое против тяжелого) (Рисунок 1B). Уровни IL6, который секретируется клетками как врожденного, так и адаптивного иммунитета, оставались неизменными между группами при стимуляции in vitro ( p = 0,1247) (рис. 1C). Что касается адаптивного иммунитета, функциональное истощение наблюдалось для лимфоцитов Th27, продуцирующих IL17A, у пациентов с тяжелой формой COVID-19 по сравнению с легкими формами и здоровыми субъектами ( p = 0,0004 и p = 0,002 соответственно) (рисунок 1D).Поразительно, что более низкая секреция IFNγ коррелировала с увеличением тяжести COVID-19 ( p <0,0001) (рис. 1E). Эта более низкая продукция IFNγ оставалась значимой даже после поправки на количество лимфоцитов ( p = 0,0183 легкое против умеренного и p = 0,0009 легкое против тяжелого) (Рисунок 1F).
Таблица 3 . Уровни цитокинов в сыворотке после неспецифической стимуляции Т-лимфоцитов и ДК у здоровых доноров и у пациентов с COVID-19 на исходном уровне.
Рисунок 1 . Уровни цитокинов в сыворотке после стимуляции in vitro клеток врожденного и адаптивного иммунитета у здоровых субъектов и у пациентов с COVID-19 с легкими, умеренными или тяжелыми симптомами. Уровни IFNα ( A : при COVID-19 легкой степени не хватает 3 баллов, при COVID-19 средней степени тяжести — 3 балла, а у пациентов с тяжелой формой COVID-19 — 6 баллов), IL6 ( C : 3, 3 , и 6 баллов соответственно), IL17 ( D ; 3, 7 и 9 баллов соответственно) и IFNγ ( E : 2, 3 и 9 баллов соответственно), а также уровни IFNα и IFNγ были скорректированы на количество моноцитов (B) и лимфоцитов (F) соответственно.Различия между группами сравнивали с тестом Краскела-Уоллиса с использованием апостериорного теста Данна .
Корреляция между продукцией IFNs после
стимуляции in vitro и осложнений, связанных с COVID-19Уровень продукции IFNα и IFNγ при стимуляции in vitro клеток врожденного и адаптивного иммунитета при поступлении и до специфического лечения прогнозировал риск осложнений ( p = 0,003 и p <0.0001 соответственно) (Рисунки 2А, В). Действительно, у пациентов с уровнем IFNα и IFNγ ниже 2,1 пг / мл и 15 МЕ / мл соответственно, как определено кривой ROC (данные не показаны), с большей вероятностью разовьются осложнения во время госпитализации ( p = 0,0098 и p = 0,0002 соответственно) (Рисунки 2B, D). Как подтверждается также многопараметрическим анализом (таблица 4), стимулированные уровни IFNγ являются независимым предиктором осложнений у пациентов с COVID-19 [ p = 0,0349 OR = 0.98 (0,962; 0,999)].
Рисунок 2 . Прогностическая ценность IFNα и IFNγ после неспецифической стимуляции клеток врожденного и адаптивного иммунитета для пациентов с COVID-19. Уровень IFNα ( A ; n = 55) и IFNγ ( C ; n = 89) различается у пациентов с осложнениями и без них и позволяет прогнозировать выживаемость без осложнений для пациентов с COVID-19. (Б, Д) . Различия между группами сравнивали с помощью теста Манна-Уитни, а для сравнения данных о выживаемости использовали лог-ранговый тест.
Таблица 4 . Многопараметрический анализ для оценки взаимосвязи между рассматриваемыми переменными на исходном уровне и осложнениями.
Эволюция уровней цитокинов в зависимости от клинического результата
Мы дополнительно оценили эволюцию продукции цитокинов и ответа на IFN во время госпитализации в умеренных и тяжелых случаях. Во время последующего наблюдения уровни нестимулированного ИЛ6 в плазме снизились между моментом поступления в больницу и последним перенесенным наблюдением для пациентов с благоприятным исходом ( p = 0.02148) (Рисунок 3A и дополнительный рисунок 1A), в то время как они оставались высокими у умерших пациентов ( p = 0,5625) (рисунок 3B и дополнительный рисунок 1B). Однако уровень IFNγ после стимуляции in vitro существенно не отличался между временем поступления в больницу и последней наблюдаемой временной точкой (рисунки 3C, D), что, вероятно, было связано с небольшим количеством пациентов в группе. . Были выбраны два индивидуальных случая, чтобы лучше продемонстрировать эволюцию продукции цитокинов в течение болезни.Первый случай привел к выздоровлению с повышенными уровнями стимулированного IFNγ в последней точке (дополнительный рисунок 1A), в то время как второй случай привел к смерти со стабильно низкими уровнями стимулированного IFNγ на протяжении всей госпитализации (дополнительный рисунок 1B).
Рисунок 3 . Эволюция уровней цитокинов у отдельных пациентов после их клинического исхода. Уровни IL6 в нестимулированной плазме ( A, B ; n = 13 и n = 6 соответственно) и уровни IFNγ после in vitro неспецифической стимуляции клеток врожденного и адаптивного иммунитета ( C, D ; n = 6 и n = 3, соответственно) сравнивались между пациентами, вылечившимися от инфекции SARS-CoV-2 (A, C) , и умершими пациентами (B , Г) .Различия между группами сравнивали с помощью знакового рангового критерия Уилкоксона для согласованных пар.
Влияние
in vitro Лечение терапевтическими молекулами на восстановление баланса цитокиновНесколько препаратов, обычно используемых для лечения пациентов с COVID-19, были протестированы на их способность восстанавливать баланс цитокинов in vitro , в частности, увеличивать продукцию IFNγ и снижать продукцию воспалительных цитокинов, сохраняя при этом постоянную секрецию регуляторных цитокинов.Хлорохин и метилпреднизолон оказались эффективными в снижении секреции всех цитокинов (рис. 4), в то время как адалимумаб снижал секрецию только IL6 и IL10. Интересно, что IFNα имел более сбалансированный эффект с сильной стимуляцией IFNγ и уменьшением воспалительного цитокина IL1β, в то время как секреция Т-регуляторного цитокина IL10 и провоспалительного цитокина (IL6) оставалась неизменной. Отдельные результаты подробно описаны в дополнительной таблице 3.
Рисунок 4 . Эффективность лечения in vitro различными препаратами, обычно используемыми при COVID-19 для модуляции экспрессии цитокинов.Уровни IFNγ (A) , IL1β (B) , IL6 (C) и IL10 (D) после предварительной обработки лекарственными средствами in vitro с последующей неспецифической стимуляцией врожденных и адаптивных иммунные клетки у 18 пациентов с COVID-19. Различия между группами сравнивали с тестом Краскела-Уоллиса с использованием апостериорного теста Данна .
Обсуждение
Мы сообщаем здесь о когорте из 101 пациента с симптоматикой инфекции COVID-19. Мы стремились выявить их специфические иммунологические профили и сопоставить их с развитием и степенью симптомов у отдельных пациентов.Наши результаты подтвердили функциональное истощение продукции типа I (NK-клетки и DC) и IFN (T-клетки) типа II у пациентов со средней и тяжелой степенью тяжести, что приводит к уклонению от врожденного и адаптивного иммунного ответа, и в соответствии с недавними исследованиями (14, 16, 18, 21, 47–49).
Хорошо известно, что врожденный иммунный ответ запускается инфицированными вирусом клетками, которые могут распознаваться рецепторами распознавания паттернов хозяина (PRR), экспрессируемыми DC, которые продуцируют различные цитокины (50), такие как IFN типа I, которые в поверните рекрут лимфоцитов и моноцитов в воспаленные участки (51–54).IFN типа I в первую очередь активирует эпителиальные клетки и снижает провоспалительную активность, опосредованную мононуклеарными макрофагами (55). IFN типа II имеют разные функции, вызывая иммунные ответы, управляемые T-хелпером 1 (Th2), а также позволяя индуцированным регуляторным T (Treg) -клеткам контролировать и регулировать иммунные ответы (56). Следовательно, SARS-CoV-2 развил несколько механизмов, чтобы ингибировать индукцию IFN типа I и передачу сигналов (57). Во время инфекции SARS-CoV-2 для успешного выведения вируса требуется как врожденный, так и адаптивный иммунный ответ, и их необходимо адекватно контролировать, чтобы минимизировать иммунопатологические повреждения (57).Оценивая ответ иммунных клеток инфицированных пациентов после стимуляции, мы демонстрируем здесь заметное снижение ответа IFN типа I и типа II от легких до тяжелых пациентов. Молекулярный механизм (ы) этого уклонения от IFN еще предстоит подтвердить, однако в нескольких исследованиях были предложены различные пути, которые могут способствовать снижению количества IFN у пациентов с тяжелой формой COVID-19, от скрытой вирусной продукции, невидимой для PPAR, до прямого синтеза структурные и неструктурные вирусные белки, которые противодействуют передаче сигналов IFN (47–49).Действительно, SARS-CoV-2 индуцировал аберрантный ответ IFN типа I в культивируемых клетках, характеризующийся отсроченным противовирусным ответом, который может обеспечивать окно для репликации вируса и неправильный набор популяций воспалительных макрофагов моноцитов (21).
Оригинальность нашей работы заключается в стимуляции TLR7 и TLR8, которые воспроизводят in vitro, вирусную инфекцию путем активации врожденной иммунной системы и продуцируют IFN типа I (58). С другой стороны, стимуляция Т-лимфоцитов анти-CD3 позволила нам количественно определить продукцию IFN типа II и оценить адаптивный иммунный ответ.Врожденное иммунное распознавание вирусной инфекции запускает противовирусный иммунный ответ остаточной геномной РНК, распознаваемой PRR, экспрессируемой в основном DC (59). У пациентов с COVID-19 средней и тяжелой степени тяжести мы наблюдали, что врожденные клетки вырабатывают меньше IFN типа I, и, следовательно, NK-клетки вырабатывают меньше IFN типа II. В соответствии с предыдущими исследованиями (21, 60), наши результаты показывают, что неконтролируемая инфекция поддерживает активацию моноцитов и макрофагов, и что регуляторные Т-лимфоциты остаются инактивированными из-за слабой продукции IFN типа II, что усиливает цитокиновый шторм и приводит к тяжелые осложнения у пациентов с COVID-19.
Это продольное исследование позволило нам сделать вывод, что функциональный анализ продукции IFN в начале госпитализации является мощным инструментом для прогнозирования клинической эволюции пациентов, инфицированных SARS-CoV-2. В то время как предыдущие исследования продемонстрировали противоположные результаты с нарушением (14, 28) или повышенным (61) ответом на IFN типа I у пациентов с тяжелой формой COVID-19, наши результаты склоняют чашу весов в сторону нарушения передачи сигналов IFN. Согласно нашему исследованию стимулированного производства IFN, а также другим исследованиям (14, 21), IFN типа I играет важную роль в активации IFN типа II и представляет собой стратегическую цель для раннего лечения пациентов с COVID-19, чтобы для уничтожения иммунного уклонения, вызванного SARS-CoV-2, и для лечения специфической иммунной дисфункции.
Однако появляется все больше доказательств того, что пациенты с тяжелой формой COVID-19 могут иметь устойчивый ответ на IFN типа I, который контрастирует с отсроченным, возможно, подавленным ответом на IFN, наблюдаемым на ранней стадии инфекции (27, 62). Ограничиваясь небольшим размером выборки из восьми и семи пациентов соответственно, Zhou et al. и Wilk et al. продемонстрировали, что многие IFN-стимулированные гены сверхэкспрессируются у пациентов с COVID-19 (62, 63).
Поскольку необходимы дополнительные исследования для дальнейшего изучения роли IFN в COVID-19, как с клинической, так и с молекулярной точки зрения, лечение IFN также остается спорным.Тем не менее, исследование in vitro на культивируемых клетках показало потенциальную пользу лечения IFNβ (48), а также недавнее клиническое исследование NCT04276688 с благоприятным исходом для IFNβ (33), в то время как другие все еще продолжаются. Два недавних ретроспективных исследования показали, что лечение IFNα может быть полезным для пациентов с COVID-19 (37, 38), однако кажется, что адекватное время введения IFN имеет решающее значение для его эффективности, поскольку раннее введение снижает смертность, в то время как позднее введение имеет противоположный эффект ( 38).
Наше исследование имеет несколько ограничений. Во-первых, это обсервационное исследование, показывающее связь между уровнями IFN I и II и тяжестью COVID-19 и исходом. Рандомизированные клинические испытания с использованием функционального анализа интерферона при поступлении для прогнозирования исхода необходимы для четкой оценки эффективности и полезности измерения интерферона в клинической практике. Во-вторых, хотя наши тесты in vitro, и несколько недавно опубликованных исследований (33, 37, 38, 48) показывают потенциал лечения IFNα для восстановления цитокинового баланса, результаты, тем не менее, должны быть подтверждены в крупных контролируемых клинических испытаниях.В-третьих, это исследование на относительно небольшом количестве пациентов, которое необходимо подтвердить в более крупных когортах. Примечательно, что в нашей когорте пациентов с COVID-19 было всего шесть смертей, что серьезно ограничивает возможности статистического анализа. В-четвертых, хотя было показано, что мужской пол, пожилой возраст и ожирение тесно связаны с повышенной смертностью пациентов с COVID-19 (10, 11), низкое количество смертей не позволило нам определить эти факторы в нашей когорте. Вместо этого мы проверили прогностическую способность возраста, пола и ИМТ в отношении осложнений, связанных с COVID-19, но, кроме уровней IL6 в плазме и стимулированных уровней IFNγ, другие переменные оставались незначимыми при многомерном анализе.
Изменчивость симптоматики лежит в основе наших клеток среди иммунных ответов, участвующих в борьбе с инфекцией COVID-19. IFNγ представляет собой прогностический биомаркер эволюции SARS-CoV-2, который можно безопасно и регулярно измерять в лаборатории с помощью QuantiFERON Monitor. Это могло бы позволить клиницистам обеспечить скорректированное лечение и медицинскую помощь в условиях эпидемии. Основываясь на наших результатах, наш функциональный тест может быть важным инструментом для прогнозирования тяжелой формы COVID-19 и руководства персонализированной терапией, направленной на восстановление иммунной системы NK и T-клеток (ингибиторы контрольных точек) и выработку IFN.
Заявление о доступности данныхНеобработанные данные, подтверждающие выводы этой статьи, будут предоставлены авторами без излишних оговорок.
Заявление об этике
Исследования с участием людей были рассмотрены и одобрены CPP SUD-OUEST et OUTRE-MER I. Пациенты / участники предоставили письменное информированное согласие на участие в этом исследовании.
Вклад авторов
CR, VB, MC, SB и BS-P разработали исследование.В.Б. и М.К. проводили эксперименты. KZ, CF, CR и MC собирали клинические данные. BS-P, CR, VB и MC проанализировали и интерпретировали данные. BS-P, CR, ED, KR, JC, EC, CI, JD и TP обеспечивали медицинский надзор. BS-P, CR и VB подготовили и отредактировали рукопись. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Это исследование было поддержано грантом Департамента приморских Альп и Национальным агентством исследований (ANR).
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим всех пациентов, участвовавших в этом исследовании, а также Элоди Малле, Жеральдин Дальмассо и Наталью Шозену за их помощь в лаборатории.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https: // www.frontiersin.org/articles/10.3389/fmed.2020.603961/full#supplementary-material
Ссылки
1. Ли Кью, Гуань Х, Ву П, Ван Х, Чжоу Л., Тонг И и др. Динамика ранней передачи новой пневмонии, инфицированной коронавирусом, в Ухане, Китай. N Engl J Med. (2020) 382: 1199–207. DOI: 10.1056 / NEJMoa2001316
PubMed Аннотация | CrossRef Полный текст | Google Scholar
2. Чжоу П., Ян Х Лу, Ван XG, Ху Б., Чжан Л., Чжан В. и др. Вспышка пневмонии, связанная с новым коронавирусом, вероятно, происхождения летучих мышей. Природа. (2020) 579: 270–3. DOI: 10.1038 / s41586-020-2951-z
PubMed Аннотация | CrossRef Полный текст | Google Scholar
3. Хуанг Ц., Ван И, Ли Х, Рен Л., Чжао Дж, Ху И и др. Клинические особенности пациентов, инфицированных новым коронавирусом 2019 г., в Ухане, Китай. Ланцет. (2020) 395: 497–506. DOI: 10.1016 / S0140-6736 (20) 30183-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
4. Гуань В.Дж., Ни З.Й., Ху И, Лян У.Х., Оу К.К., Хе Дж. Х и др.Клиническая характеристика коронавирусной болезни 2019 в Китае. N Engl J Med . (2020) 382: 1708–20. DOI: 10.1056 / NEJMoa2002032
CrossRef Полный текст | Google Scholar
5. Jin YH, Cai L, Cheng ZS, Cheng H, Deng T., Fan YP и др. Краткое руководство по диагностике и лечению пневмонии, инфицированной новым коронавирусом 2019 (2019-nCoV) (стандартная версия). Mil Med Res. (2020) 7: 1–23. DOI: 10.1186 / s40779-020-0233-6
CrossRef Полный текст
6.Wu Z, McGoogan JM. Характеристики и важные уроки вспышки коронавирусного заболевания 2019 г. (COVID-19) в Китае: краткое изложение отчета Китайского центра по контролю и профилактике заболеваний о 72314 случаях. J Am Med Assoc. (2020) 323: 1239–42. DOI: 10.1001 / jama.2020.2648
PubMed Аннотация | CrossRef Полный текст | Google Scholar
8. Хэнли Б., Нареш К.Н., Руфосс С., Николсон А.Г., Вейр Дж., Кук Г.С. и др. Гистопатологические данные и вирусный тропизм у британских пациентов с тяжелым смертельным исходом COVID-19: посмертное исследование. Ланцетный микроб. (2020) 1: e245–53. DOI: 10.1016 / S2666-5247 (20) 30115-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
9. Лю К., Фанг Й.Й., Дэн Й., Лю В., Ван М.Ф., Ма Дж. П. и др. Клиническая характеристика случаев нового коронавируса в больницах третичного уровня провинции Хубэй. Chin Med J. (2020) 133: 1025–31. DOI: 10.1097 / CM9.0000000000000744
CrossRef Полный текст | Google Scholar
10. Гун Дж, Оу Дж, Цю Х, Цзе Й, Чен Й, Юань Л. и др.Инструмент для раннего прогнозирования тяжелой коронавирусной болезни 2019 (COVID-19): многоцентровое исследование с использованием номограммы риска в Ухане и Гуандуне, Китай. Клин Инфекция Дис . (2020) 71: 833–40. DOI: 10.1093 / cid / ciaa443
PubMed Аннотация | CrossRef Полный текст | Google Scholar
11. Чен X, Лю З. Раннее прогнозирование риска смертности среди пациентов с тяжелой формой COVID-19 с использованием машинного обучения. Int J Epidemiol . (2020) 42: dyaa171. DOI: 10.1093 / ije / dyaa171
CrossRef Полный текст | Google Scholar
12.Карвелли Дж., Демария О., Вели Ф., Батиста Л., Бенмансур, Северная Каролина, Фарес Дж. И др. Ассоциация воспаления COVID-19 с активацией оси C5a – C5aR1. Природа . (2020) 588: 146–50. DOI: 10.1038 / s41586-020-2600-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
13. Вабрет Н., Бриттон Дж. Дж., Грубер С., Хегде С., Ким Дж., Куксин М. и др. Иммунология COVID-19: современное состояние науки. Иммунитет . (2020) 52: 910–41.
PubMed Аннотация | Google Scholar
14.Hadjadj J, Yatim N, Barnabei L, Corneau A, Boussier J, Pere H и др. Нарушение активности интерферона I типа и обострение воспалительных реакций у пациентов с тяжелым Covid-19. Наука (80-) . (2020) 369: 718–24. DOI: 10.1126 / science.abc6027
PubMed Аннотация | CrossRef Полный текст | Google Scholar
15. Фей Дж, Фу Л, Ли И, Сян Х. Х, Сян И, Ли М-Д и др. Уменьшение количества лимфоцитов на ранней стадии увеличивает тяжесть и риск смерти пациентов с COVID-19: когортное исследование на базе больниц. MedRxiv . (2020). DOI: 10.1101 / 2020.04.02.20050955
CrossRef Полный текст | Google Scholar
17. Johannessen LE, Pedersen NM, Pedersen KW, Madshus IH, Stang E. Активация рецептора эпидермального фактора роста (EGF) индуцирует образование ямок, покрытых клатрином, содержащих рецептор EGF и Grb2. Mol Cell Biol. (2006) 26: 389–401. DOI: 10.1128 / MCB.26.2.389-401.2006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
19.Рагаб Д., Салах Элдин Х., Тейма М., Хаттаб Р., Салем Р. Цитокиновый шторм COVID-19: что мы знаем до сих пор. Front Immunol. (2020) 11: 1–4. DOI: 10.3389 / fimmu.2020.01446
PubMed Аннотация | CrossRef Полный текст
20. Chen G, Wu D, Guo W., Cao Y, Huang D, Wang H, et al. Клинические и иммунологические особенности при тяжелой и умеренной коронавирусной болезни 2019. J Clin Invest . (2020) 130: 2620–9. DOI: 10.1101 / 2020.02.16.20023903
PubMed Аннотация | CrossRef Полный текст | Google Scholar
21.Бланко-Мело Д., Нильссон-Пайант Б. Э., Лю В. К., Уль С., Хоугланд Д., Мёллер Р. и др. Несбалансированная реакция хозяина на SARS-CoV-2 способствует развитию COVID-19. Ячейка . (2020) 181: 1036–45. DOI: 10.1016 / j.cell.2020.04.026
PubMed Аннотация | CrossRef Полный текст | Google Scholar
23. Du SQ, Yuan W. Математическое моделирование взаимодействия между врожденными и адаптивными иммунными ответами при COVID-19 и их значение для вирусного патогенеза. J Med Virol . (2020) 92: 1615–28.DOI: 10.1002 / jmv.25866
PubMed Аннотация | CrossRef Полный текст | Google Scholar
24. Вивье Э., Томаселло Э., Баратин М., Вальцер Т., Уголини С. Функции естественных клеток-киллеров. Nat Immunol. (2008) 9: 503–10. DOI: 10.1038 / ni1582
CrossRef Полный текст | Google Scholar
25. Бонжардим CA. Интерфероны (IFN) являются ключевыми цитокинами как в врожденном, так и в адаптивном противовирусном иммунном ответе, а вирусы противодействуют действию IFN. Microbes Infect. (2005) 7: 569–78. DOI: 10.1016 / j.micinf.2005.02.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
26. Жесткий DF. Интерферон I типа как связующее звено между врожденным и адаптивным иммунитетом посредством стимуляции дендритных клеток. Лейк Лимфома. (2004) 45: 257–64. DOI: 10.1080 / 104281
- 00149368
- •
Норовирус человека (HuNoV) обладал высокой чувствительностью к IFN типа I, II и III лечение.
- •
Сверхэкспрессия важных человеческих ISG идентифицировала IRF-1, RIG-I и MDA5 как сильные анти-норовирусные эффекторы.
- •
IFN типа I / III координированно индуцируют IRF-1, RIG-I и MDA5, тогда как IFN типа II преимущественно индуцируют IRF-1 для борьбы с HuNoV.
- •
IFN типа II в сочетании с IFN типа I или III оказывал синергетический противовирусный эффект против репликации РНК HuNoV.
- •
Репликация HuNoV не влияла на сигнальный путь IFNs.
- 1. Котенко С.В., Галлахер Г., Баурин В.В., Льюис-Антес А., Шен М. и др. (2003) IFN-lambdas опосредуют противовирусную защиту через отдельный рецепторный комплекс цитокинов класса II. Нат Иммунол 4: 69–77.
- 2. Шеппард П., Киндсфогель В., Сюй В., Хендерсон К., Шлюцмайер С. и др. (2003) IL-28, IL-29 и их рецептор цитокинов класса II IL-28R. Нат Иммунол 4: 63–68.
- 3. Pagliaccetti NE, Robek MD (2010) Интерферон-λ при инфекции и терапии ВГС.Вирусы 2: 1589–1602.
- 4. Анк Н., Иверсен МБ, Бартольди С., Стахели П., Хартманн Р. и др. (2008) Важная роль интерферона типа III (IFN-lambda / IL-28) в TLR-индуцированной противовирусной активности. J Immunol 180: 2474–2485.
- 5. Робек MD, Boyd BS, Chisari FV (2005) Лямбда-интерферон ингибирует репликацию вирусов гепатита B и C. J Virol 79: 3851–3854.
- 6. Хоу В., Ван Х, Е Л., Чжоу Л., Ян З. К. и др. (2009) Лямбда-интерферон подавляет инфицирование макрофагов вирусом иммунодефицита человека 1 типа.J Virol 83: 3834–3842.
- 7. Анк Н., Вест Х., Бартольди К., Эрикссон К., Томсен А. Р. и др. (2006) Лямбда-интерферон (IFN-lambda), IFN типа III, индуцируется вирусами и IFN и проявляет мощную противовирусную активность против некоторых вирусных инфекций in vivo. J Virol 80: 4501–4509.
- 8. Миллер Д.М., Клучер К.М., Фриман Дж. А., Хаусман Д. Ф., Фонтана Д. и др. (2009) Интерферон-лямбда как потенциальное новое лекарство от гепатита С. Ann N Y Acad Sci 1182: 80–87.
- 9. Ли М., Лю X, Чжоу Ю., Су С.Б. (2009) Интерферон-лямбды: модуляторы антивирусных, противоопухолевых и иммунных ответов. J Leukoc Biol 86: 23–32.
- 10. Чжан Л., Джилг Н., Шао RX, Лин В., Фуско Д. Н. и др. (2011) IL28B подавляет репликацию вируса гепатита C через путь JAK-STAT. J Hepatol 55: 289–298.
- 11. Ge D, Fellay J, Thompson AJ, Simon JS, Shianna KV и др. (2009) Генетическая изменчивость IL28B предсказывает вирусный клиренс, вызванный лечением гепатита С.Природа 461: 399–401.
- 12. Thomas DL, Thio CL, Martin MP, Qi Y, Ge D и др. (2009) Генетическая изменчивость IL28B и спонтанное избавление от вируса гепатита С. Природа 461: 798–801.
- 13. Танака Ю., Нисида Н., Сугияма М., Куросаки М., Мацуура К. и др. (2009) Общегеномная ассоциация IL28B с ответом на терапию пегилированным интерфероном-альфа и рибавирином хронического гепатита C. Nat Genet 41: 1105–1109.
- 14. Суппиа В., Молдавский М., Аленстиэль Г., Берг Т., Велтман М. и др.(2009) IL28B связан с ответом на терапию интерфероном-альфа и рибавирином при хроническом гепатите С. Нат Генет 41: 1100–1104.
- 15. Уилли Р.Л., Смит Д.Х., Ласки Л.А., Теодор Т.С., Эрл П.Л. и др. (1988) Мутагенез in vitro идентифицирует область внутри гена оболочки вируса иммунодефицита человека, которая имеет решающее значение для инфекционности. J Virol 62: 139–147.
- 16. Хо В.З., Лиой Дж., Сонг Л., Кутилли Дж. Р., Полин Р.А. и др. (1992) Инфекция макрофагов, происходящих из моноцитов пуповинной крови, вирусом иммунодефицита человека 1 типа.J Virol 66: 573–579.
- 17. Sadler AJ, Williams BR (2008) Интерферон-индуцируемые противовирусные эффекторы. Nat Rev Immunol 8: 559–568.
- 18. Каваи Т., Акира С. (2010) Роль рецепторов распознавания образов в врожденном иммунитете: обновленная информация о Toll-подобных рецепторах. Нат Иммунол 11: 373–384.
- 19. Ank N, Paludan SR (2009) IFN типа III: новые уровни сложности врожденного противовирусного иммунитета. Биофакторы 35: 82–87.
- 20. Iversen MB, Ank N, Melchjorsen J, Paludan SR (2010) Экспрессия интерферона типа III (IFN) в слизистой оболочке влагалища опосредуется в основном дендритными клетками и проявляет более сильную зависимость от NF-kappaB, чем IFN типа I.J Virol 84: 4579–4586.
- 21. Megjugorac NJ, Gallagher GE, Gallagher G (2010) IL-4 усиливает продукцию IFN-lambda1 (IL-29) плазматическими DC посредством секреции IL-1Ra моноцитами. Кровь 115: 4185–4190.
- 22. Акира С., Такеда К. (2004) Передача сигналов толл-подобного рецептора. Nat Rev Immunol 4: 499–511.
- 23. Alexopoulou L, Holt AC, Medzhitov R, Flavell RA (2001) Распознавание двухцепочечной РНК и активация NF-kappaB Toll-подобным рецептором 3.Природа 413: 732–738.
- 24. Чжоу Ю., Ван Х, Лю М., Ху Ц., Сун Л. и др. (2010) Критическая функция toll-подобного рецептора-3 в индукции активности вируса иммунодефицита человека в макрофагах. Иммунология 131: 40–49.
- 25. Barnes BJ, Moore PA, Pitha PM (2001) Вирус-специфическая активация нового фактора регуляции интерферона, IRF-5, приводит к индукции отдельных генов интерферона альфа. J Biol Chem 276: 23382–23390.
- 26.Marie I, Durbin JE, Levy DE (1998) Дифференциальная вирусная индукция отдельных генов интерферона-альфа посредством положительной обратной связи через фактор регуляции интерферона-7. Embo J 17: 6660–6669.
- 27. Au WC, Pitha PM (2001) Рекрутирование множества факторов регуляции интерферона и гистонацетилтрансферазы на транскрипционно активные промоторы интерферона а. J Biol Chem 276: 41629–41637.
- 28. Barnes BJ, Kellum MJ, Field AE, Pitha PM (2002) Множественные регуляторные домены IRF-5 контролируют активацию, клеточную локализацию и индукцию хемокинов, которые опосредуют рекрутирование Т-лимфоцитов.Mol Cell Biol 22: 5721–5740.
- 29. Pagliaccetti NE, Robek MD (2010) Интерферон-лямбда в инфекции и терапии ВГС.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
29. Бойер-Суавет С., Кремони М., Дюпейрат Т., Зорзи К., Брглез В., Бензакен С. и др.Функциональный иммунный анализ с использованием гамма-интерферона может предсказать инфекционные события в терминальной стадии заболевания почек. Clin Chim Acta. (2019) 502: 287–92. DOI: 10.1016 / j.cca.2019.11.018
PubMed Аннотация | CrossRef Полный текст | Google Scholar
31. Хорнусс Д., Лаубнер К., Монастерио С., Тимме Р., Вагнер Д. [COVID-19-ассоциированная пневмония, несмотря на неоднократные отрицательные результаты ПЦР-анализа мазков из ротоглотки]. Dtsch Med Wochenschr . (2020) 145: 844–9. DOI: 10.1055 / a-1170-6061
PubMed Аннотация | CrossRef Полный текст | Google Scholar
32.Bouaziz JD, Duong T, Jachiet M, Velter C, Lestang P, Cassius C и др. Сосудистые кожные симптомы при COVID-19: французское обсервационное исследование. J Eur Acad Dermatol Venereol . (2020) 34: e451–2. DOI: 10.1111 / jdv.16544
PubMed Аннотация | CrossRef Полный текст | Google Scholar
33. Hung IFN, Lung KC, Tso EYK, Liu R, Chung TWH, Chu MY, et al. Тройная комбинация интерферона бета-1b, лопинавира – ритонавира и рибавирина в лечении пациентов, госпитализированных с COVID-19: открытое рандомизированное исследование фазы 2. Ланцет. (2020) 395: 1695–704. DOI: 10.1016 / S0140-6736 (20) 31042-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
34. Виджайваргия П., Эскер Гарригос З., Кастильо Алмейда, NE, Гуррам ПР, Стивенс Р.В., Разонабл Р.Р. Рекомендации по лечению COVID-19: критический обзор доказательств (или их отсутствия). Mayo Clin Proc. (2020) 95: 1454–66. DOI: 10.1016 / j.mayocp.2020.04.027
PubMed Аннотация | CrossRef Полный текст | Google Scholar
35.Коллективная группа RECOVERY. Дексаметазон у госпитализированных пациентов с Covid-19 — предварительное сообщение. N Engl J Med . (2020). [Epub перед печатью].
Google Scholar
37. Чжоу К., Чен В., Шеннон С.П., Вэй XS, Сян Х, Ван Х и др. Лечение COVID-19 интерфероном-α2b. Front Immunol. (2020) 11: 1–6. DOI: 10.3389 / fimmu.2020.615275
CrossRef Полный текст | Google Scholar
38. Ван Н, Чжань И, Чжу Л., Хоу З, Лю Ф, Сун П. и др.Ретроспективное многоцентровое когортное исследование показывает, что ранняя терапия интерфероном связана с благоприятным клиническим ответом у пациентов с COVID-19. Клеточный микроб-хозяин. (2020) 28: 455–64.e2. DOI: 10.1016 / j.chom.2020.07.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
39. Вирсинга В.Дж., Родос А., Ченг А.С., Пикок С.Дж., Прескотт ХК. Патофизиология, передача, диагностика и лечение коронавирусной болезни 2019 (COVID-19): обзор. JAMA — J Am Med Assoc. (2020) 324: 782–93. DOI: 10.1001 / jama.2020.12839
PubMed Аннотация | CrossRef Полный текст | Google Scholar
40. Ризк Дж. Г., Калантар-Заде К., Мехра М. Р., Лави С. Дж., Ризк Й., Фортал Д. Н.. Фармако-иммуномодулирующая терапия при COVID-19. Наркотики. (2020) 80: 1267–92. DOI: 10.1007 / s40265-020-01367-z
CrossRef Полный текст | Google Scholar
41. Сабадош Б., Абу-Ганем Ю., Грант М., Чой Дж., Бекс А., Поулз Т. Клинические характеристики и исход для четырех больных раком, инфицированных SARS-CoV-2, которых лечили ингибиторами иммунных контрольных точек. Eur Urol. (2020) 78: 276–80. DOI: 10.1016 / j.eururo.2020.05.024
PubMed Аннотация | CrossRef Полный текст | Google Scholar
43. Саадун Д., Розенцвайг М., Джоли Ф., Сикс А., Каррат Ф., Тибо В. и др. Регуляторные Т-клеточные ответы на низкие дозы интерлейкина-2 при ВГС-индуцированном васкулите. N Engl J Med. (2011) 365: 2067–77. DOI: 10.1056 / NEJMoa1105143
PubMed Аннотация | CrossRef Полный текст | Google Scholar
44. Патинот С., Каррум Н.Б., Моарбесс Г., Чирнат Н., Кассаб И., Боннет П.А. и др.Лиганды-агонисты и антагонисты толл-подобных рецепторов 7 и 8: оригинальные инструменты для терапевтических целей. Eur J Med Chem. (2020) 193: 112238. DOI: 10.1016 / j.ejmech.2020.112238
PubMed Аннотация | CrossRef Полный текст | Google Scholar
45. Крозат К., Вивье Э., Далод М. Перекрестное взаимодействие между компонентами врожденной иммунной системы: повышение антимикробной защиты и предотвращение иммунопатологий. Immunol Rev. (2009) 227: 129–49. DOI: 10.1111 / j.1600-065X.2008.00736.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
46. Zheng M, Gao Y, Wang G, Song G, Liu S, Sun D и др. Функциональное истощение противовирусных лимфоцитов у пациентов с COVID-19. Cell Mol Immunol. (2020) 17: 533–5. DOI: 10.1038 / s41423-020-0402-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
48. Лэй X, Донг X, Ма Р, Ван В., Сяо Х, Тиан З. и др. Активация и уклонение от ответа на интерферон I типа SARS-CoV-2. Nat Commun. (2020) 11: 1–12. DOI: 10.1038 / s41467-020-17665-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
52. Ито Т., Ван Ю.Х., Лю Ю.Дж. Плазмацитоидные предшественники дендритных клеток / клетки, продуцирующие интерферон I типа, обнаруживают вирусную инфекцию с помощью Toll-подобного рецептора (TLR) 7 и TLR9. Springer Semin Immunopathol. (2005) 26: 221–9. DOI: 10.1007 / s00281-004-0180-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
53.Теофилопулос А.Н., Баккала Р., Бейтлер Б., Коно Д.Х. Интерфероны I типа (A / B) при иммунитете и аутоиммунитете. Annu Rev Immunol. (2005) 23: 307–35. DOI: 10.1146 / annurev.immunol.23.021704.115843
CrossRef Полный текст | Google Scholar
54. Фор Э., Пуасси Дж., Гоффард А., Фурнье С., Кипнис Э., Титекат М. и др. Отчетливый иммунный ответ у двух пациентов, инфицированных БВРС-КоВ: можем ли мы перейти от скамейки к постели? PLoS ONE. (2014) 9: e88716. DOI: 10.1371 / journal.pone.0088716
PubMed Аннотация | CrossRef Полный текст | Google Scholar
55. Дэвидсон С., МакКейб Т.М., Кротта С., Гад Х.Х., Хессель Е.М., Бинке С. и др. IFN λ является мощным противогриппозным лекарственным средством без побочных воспалительных эффектов лечения IFN α. EMBO Mol Med. (2016) 8: 1099–112. DOI: 10.15252 / emmm.201606413
PubMed Аннотация | CrossRef Полный текст | Google Scholar
57. Лагунас-Рангель Ф.А., Чавес-Валенсия V. Высокое соотношение IL-6 / IFN-γ может быть связано с тяжелым заболеванием у пациентов с COVID-19. J Med Virol . (2020) 92: 1789–90. DOI: 10.1002 / jmv.25900
PubMed Аннотация | CrossRef Полный текст | Google Scholar
58. Суд С., Кандалл Д., Ю. Л., Миямасу М., Бойл Дж. С., Онг С.И. и др. Новый биомаркер иммунной функции и первоначальный опыт в популяции трансплантатов. Трансплантация. (2014) 97: 50–1. DOI: 10.1097 / TP.0000000000000078
PubMed Аннотация | CrossRef Полный текст | Google Scholar
60. Морейра-Тейшейра Л., Майер-Барбер К., Шер А., О’Гарра А.Интерфероны I типа при туберкулезе: враг, а иногда и друг. J Exp Med. (2018) 215: 1273–85. DOI: 10.1084 / jem.20180325
PubMed Аннотация | CrossRef Полный текст | Google Scholar
61. Лукас С., Вонг П., Кляйн Дж., Кастро ТБР, Сильва Дж., Сундарам М. и др. Продольный анализ выявляет иммунологические перебои в работе при тяжелой форме COVID-19. Природа. (2020) 584: 463–9. DOI: 10.1038 / s41586-020-2588-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
62.Zhou Z, Ren L, Zhang L, Zhong J, Xiao Y, Jia Z и др. Повышенный врожденный иммунный ответ в дыхательных путях у пациентов с COVID-19. Клеточный микроб-хозяин. (2020) 27: 883–90.e2. DOI: 10.1016 / j.chom.2020.04.017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
63. Вилк А.Дж., Рустаги А., Чжао Н.К., Роке Дж., Мартинес-Колон Г.Дж., МакКечни Дж.Л. и др. Одноклеточный атлас периферического иммунного ответа у пациентов с тяжелой формой COVID-19. Nat Med. (2020) 26: 1070–6.DOI: 10.1038 / s41591-020-0944-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Высокий уровень циркулирующих интерферонов типа I, типа II и типа III ассоциируется с отчетливыми клиническими признаками активной системной красной волчанки | Arthritis Research & Therapy
Здесь мы представляем первое сравнительное исследование функциональной активности IFN I типа, измеренной in vitro с помощью анализа репортерных клеток WISH, метода, часто используемого в качестве золотого стандарта для оценки сигнатуры IFN при СКВ, и измерения циркулирующего типа I , IFN типа II и типа III.Наше исследование уникально, поскольку мы измерили все три типа ИФН в большой и очень хорошо охарактеризованной когорте СКВ. Этот подход позволил нам определить, как разные подтипы IFN связаны друг с другом и с клиническими подмножествами СКВ.
Мы обнаружили, что активность IFN типа I коррелирует с циркулирующими уровнями IFN типа I и типа II, но слабо с IFN типа III, который, насколько нам известно, ранее не наблюдался. Было известно, что уровни IFN-γ увеличиваются за годы до постановки диагноза СКВ.Наблюдаемая корреляция между активностью IFN и IFN-γ предполагает, что роль IFN-γ может быть столь же важной, как IFN типа I при СКВ. Следовательно, IFN-γ также может быть важной мишенью для терапевтических стратегий. Кроме того, многомерные модели номинальной регрессии позволили нам идентифицировать фенотипические подмножества СКВ, связанные с высокими уровнями различных подтипов ИФН.
Мы сообщаем о новом наблюдении, согласно которому активный LN связан как с высокой активностью IFN, так и с высокими уровнями циркулирующего IFN-γ.Среди случаев волчаночного нефрита (активного или перенесенного) мы наблюдали долю пациентов с перекрывающимся повышением активности IFN, IFN-γ и сывороточного IFN-α, как сообщалось ранее [10, 32]. Таким образом, наши результаты расширяют предыдущие знания о том, что почечные обострения связаны с комбинированной сигнатурой генов IFN-α и IFN-γ [32, 33]. Помимо IFN-γ, другие типы IFN, по-видимому, имеют большое значение. Исследование азиатского населения показало, что высокие уровни IFN-λ1 также связаны с LN [34]. Мы не смогли подтвердить эту связь, возможно, из-за того, что некоторые пациенты уже проходили индукционную терапию при наборе; тем не менее, в нашем более раннем исследовании мы обнаружили, что высокие уровни IFN-λ1 были связаны с плохими почечными исходами [11].Интересно, что в нашей когорте высокие уровни IFN-α в сыворотке были связаны с лучшим сохранением функции почек, и это наблюдение осталось после корректировки по возрасту [11].
Артрит с СКВ был связан как с высокой активностью IFN типа I, так и с высоким IFN-γ. Ранее мы сообщали, что артрит был связан с активацией IP-10 / CXCL-10 [11]. В целом, наши наблюдения предполагают, что артрит при СКВ связан с несколькими цитокинами в путях ИФН [11].
Как и другие исследователи, наши данные демонстрируют, что поражение слизистых и кожных покровов связано с высокой активностью IFN типа I in vitro и с высокими уровнями циркулирующего IFN-α [8, 35], что позволяет предположить, что эта подгруппа может получить пользу от блокаторов IFN-α. [20].
Мы также наблюдали, что образцы аутоантител связаны с различными подтипами IFN.
Активность IFN типа I соответствует большинству субспецифичностей ANA, как сообщалось ранее [8, 31, 32, 35]. В многофакторном анализе мы выявили новую ассоциацию, согласно которой высокий уровень IFN-γ наиболее сильно связан с анти-Ro60. Мы подтверждаем предыдущие данные об IFN-α, который, как известно, связан с анти-Ro52 и анти-La [8].
В нескольких исследованиях изучалась взаимосвязь между ИФН и сосудистыми исходами при СКВ [36].Более ранние сообщения из более широких этнических групп наблюдали ассоциацию aPL и высокой активности IFN типа I в определенных группах предков [37]. В нынешней когорте, в основном кавказской, многофакторный анализ продемонстрировал более низкую вероятность сосудистых событий, назначение АФЛ и варфарина для пациентов с изолированным высоким ИФН-альфа. Кроме того, в группе с высоким уровнем IFN-γ частота сосудистых событий была несколько ниже, поскольку назначение антикоагулянтов было менее распространенным. Высокий уровень IFN-λ1 был связан с антинуклеосомными аутоантителами и численно, но не значительно, с более высокой частотой aPL, сосудистых событий и использования варфарина.Наши результаты показывают, что в этой европейской популяции IFN типа I и типа II в первую очередь связаны с аутореактивностью с внутриклеточными аутоантигенами, но не с aPL, которые нацелены на неядерные структуры, такие как белки плазмы и фосфолипиды мембран [30]. Насколько нам известно, ранее не сообщалось о связи между сердечно-сосудистыми заболеваниями и уровнями периферических уровней ИФН типа II и III.
Известно, что высокая активность IFN I типа связана с гематологическими проявлениями [35].В нашем отчете добавлена новая информация о том, что гематологические проявления также связаны с повышенным уровнем IFN-γ. Как и ожидалось, мы подтверждаем, что низкие уровни комплемента следуют за высокой активностью IFN типа I, а также за высокими уровнями IFN-α и IFN-γ [2, 9, 28, 29].
Значительная часть более ранних исследователей полагалась на оценки активации IFN-гена и их связи с активностью заболевания СКВ [32, 37, 38]. Однако каждый исследователь предпочел измерить различные гены, регулируемые IFN, и поэтому исследования трудно сравнивать.Еще одним осложняющим фактором при оценке сигнатуры IFN является то, что одни и те же гены могут регулироваться несколькими типами IFN. Наши результаты показывают, что пациенты с активной СКВ имеют высокий уровень циркулирующих ИФН типа I, типа II и типа III и что поражение различных органов, по-видимому, связано с различными типами ИФН. Представленные данные предполагают, что измерение уровней циркулирующего IFN более информативно, чем оценка сигнатуры IFN, и может иметь значение для выявления пациентов, которым анти-IFN терапия при СКВ будет полезна.
Мы демонстрируем, что несколько основных клинических проявлений связаны с доминирующим типом IFN, но все же некоторые пациенты имеют другие типы IFN с повышенной регуляцией. Было бы интересно, если бы наши наблюдения могли быть подтверждены в других когортах. В клинической практике было бы важно измерить все три типа ИФН и определить доминирующий, чтобы адаптировать направленную терапию. Кроме того, наши результаты показывают, что соединения, блокирующие несколько путей IFN в клетке, будут более многообещающим терапевтическим подходом, чем моноклональные антитела, блокирующие один циркулирующий IFN или его рецептор [20, 39].
Активация IFN-регулируемых путей I типа считается ключевым механизмом в патогенезе СКВ [40], но новые данные демонстрируют, что важную роль также играют IFN типа II и ось цитокинов Th27 [9, 11, 41]. Фаза 2 испытаний устекинумаба, антител, блокирующих путь IL-12 / IL-23, при СКВ была недавно опубликована с положительными и многообещающими результатами [42]. Хороший ответ на устекинумаб был связан со снижением уровней IFN-γ, в то время как показатели подписи IFN I типа не снижались.Ранее мы сообщали, что высокие уровни IL-23 и IL-17 связаны с резистентными к лечению ЛУ с плохими результатами [10, 36, 37]. В целом, накопленные данные подтверждают роль IFN-γ и оси Th27 в SLE, предполагая, что при тяжелых проявлениях, включая LN, могут помочь агенты, блокирующие IFN-γ и / или Th27.
IFN-β является важным членом семейства IFN типа I. К сожалению, мы не измерили циркулирующие уровни IFN-β. Возможно, что некоторые расхождения в наших результатах между анализом IFN функционального типа I и ELISA IFN-α связаны с тем, что функциональный анализ также измеряет IFN-β.Хотя это могло быть объяснением, было показано, что IFN-α является основным циркулирующим IFN типа I в большинстве сывороток при СКВ [12]. Таким образом, представляется вероятным, что функциональный анализ более чувствителен, чем ELISA, для общего измерения IFN-α [14].
Мы наблюдали, что пациенты с высокой активностью IFN имели немного более высокие OR для доз преднизолона более 10 мг, но не наблюдали никаких других ассоциаций между уровнями IFN и иммуномодулирующей терапией. Дизайн представленного исследования был перекрестным, и в него были включены пациенты с сильно различающимися показателями активности заболевания, а также различные виды лечения.Этот факт мог ограничить нашу возможность проанализировать, как различные методы лечения могут влиять на уровни IFN. Проспективное лонгитюдное исследование было бы лучшим вариантом для изучения этих вопросов.
Путь интерферона | Thermo Fisher Scientific
И рецептор IFN типа I, и рецептор IFN типа II имеют многоцепочечные структуры, которые состоят по крайней мере из двух отдельных субъединиц: IFNAR1 и IFNAR2 для рецептора IFN типа I и IFNGR1 и IFNGR2 для Рецептор IFN типа II.Каждая из этих субъединиц рецептора взаимодействует с членом семейства JAK (Janus Activated Kinase). В случае рецептора IFN типа I субъединица IFNAR1 конститутивно связана с TYK2 (тирозинкиназа-2), тогда как IFNAR2 связана с JAK1 [5-7]. В случае рецептора IFN типа II субъединица IFNGR1 ассоциирована с JAK1, тогда как IFNGR2 конститутивно ассоциирована с JAK2 [6]. Начальным этапом передачи сигналов IFN-IFN типа I и II является активация этих рецептор-ассоциированных JAK, которая происходит в ответ на лиганд-зависимую перестройку и димеризацию субъединиц рецептора с последующим аутофосфорилированием и активацией. ассоциированных JAK.Связывание IFN-Alpha или других IFN типа I с рецептором IFN типа I приводит к быстрому аутофосфорилированию и активации связанных с рецептором JAK TYK2 и JAK1 [4,8], которые, в свою очередь, регулируют фосфорилирование и активацию STAT. . STATs, которые активируются в ответ на IFN типа I, включают STAT1, STAT2, STAT3 и STAT5. Активация таких STAT является обычным ответом на разные IFN типа I, что согласуется с тем, что все эти IFN связываются с одним и тем же рецептором и, таким образом, активируют общий путь, который включает одни и те же JAK, TYK2 и JAK1.STAT4 и STAT6 также могут активироваться IFN-альфа, но такая активация, по-видимому, ограничена определенными типами клеток, такими как эндотелиальные клетки или клетки лимфоидного происхождения [9,10]. После фосфорилирования JAK активированные STAT образуют гомодимеры или гетеродимеры, которые перемещаются в ядро и инициируют транскрипцию путем связывания определенных сайтов в промоторах ISG (IFN-стимулированных генов) [6]. Важным транскрипционным комплексом, который индуцируется IFN типа I, является комплекс ISGF3 (ISG Factor-3).Зрелый комплекс ISGF3 состоит из фосфорилированных (активированных) форм STAT1 и STAT2 вместе с IRF9, который не подвергается фосфорилированию тирозина [11,13-15]. Этот комплекс является единственным комплексом, который связывает определенные элементы, известные как ISRE (IFN-стимулированные элементы ответа), которые присутствуют в промоторах определенных ISG, тем самым инициируя их транскрипцию. Другие комплексы STAT, которые индуцируются IFN типа I, включают гомодимеры STAT1 – STAT1, STAT3 – STAT3, STAT4 – STAT4, STAT5 – STAT5 и STAT6 – STAT6, а также STAT1 – STAT2, STAT1 – STAT3, STAT1 – STAT4, STAT1– Гетеродимеры STAT5, STAT2 – STAT3 и STAT5 – STAT6 [11,12].Такие IFN-индуцированные комплексы связывают другой тип элемента, известный как элемент GAS (IFN-Gamma-Activated Site), который присутствует в промоторе ISG [11]. Из сотен известных ISG некоторые имеют только ISRE или только элементы GAS в своих промоторах, тогда как другие имеют оба элемента; следовательно, комбинации различных STAT-содержащих комплексов могут потребоваться для оптимальной активации транскрипции конкретного гена. Транскрипция IFN-зависимых генов типа II (IFN-Gamma) регулируется элементами GAS, и STAT1 является наиболее важным фактором транскрипции, активируемым IFN-Gamma, для регуляции этих транскрипционных ответов.После взаимодействия IFN-рецептора II типа с помощью IFN-Gamma, JAK1 и JAK2 активируются и регулируют нижестоящее фосфорилирование STAT1 по остатку тирозина в положении Tyr701 [11]. Фосфорилированная тирозином форма STAT1 образует гомодимеры, которые перемещаются в ядро и связывают элементы GAS, которые присутствуют в промоторах регулируемых IFN-гамма генов. IFN-Gamma-активированные JAK также регулируют через еще неизвестные промежуточные соединения активацию каталитической субъединицы (p110) PI3K. Активация PI3K в конечном итоге приводит к последующей активации PKC-Delta (Protein Kinase-C-Delta), которая, в свою очередь, регулирует фосфорилирование STAT1 на Ser727.Фосфорилирование серина не является существенным для транслокации STAT1 в ядро или для связывания STAT1 с ДНК, но необходимо для полной активации транскрипции. Фосфорилированный STAT1 гомодимеризуется с образованием комплекса GAF-AAF, который перемещается в ядро и связывается с элементом GAS, присутствующим в большинстве генов, индуцируемых IFN-Gamma. В отличие от IFN типа I, IFN-Gamma не индуцирует образование комплексов ISGF3 и, таким образом, не может индуцировать транскрипцию генов, которые имеют только ISRE в своем промоторе.Подобно IFN-Gamma, передача сигналов IFN-Alpha и IFN-Beta также может приводить к образованию комплекса GAF-AAF и его связыванию с регуляторным элементом GAS. Три недавно идентифицированных белка IFN-Lambda (также называемые IL-28A, IL-28B и IL-29) связываются с гетеродимерной субъединицей рецептора цитокинов класса II IFN-LamdaR1 (также называемой CRF2-12) и второй субъединицей IL- 10R2 (CRF2-4), который служит второй цепью IL-10R. Сшивание IFN-лямбда приводит к активации как STAT1, так и STAT2, что приводит к образованию комплексов ISGF3 и GAF-AAF.Эти недавно описанные цитокины функционально подобны IFN типа I, поскольку их синтез индуцируется вирусной инфекцией или двухцепочечной РНК. Они делают клетки устойчивыми к вирусной инфекции и активируют те же внутриклеточные сигнальные пути, что и IFN типа I. Семейство IFN-лямбда представляет собой интересную эволюционную связь между IFN типа I и семейством IL-10, но они структурно более тесно связаны с IFN типа I. IFN-Lambda также активирует STAT3 и STAT5, которые более характерно связаны с передачей сигналов цитокинами, родственными IL-10 и IL-10 [3].
Другой сигнальный каскад, регулируемый IFN, — это путь MAPK (Mitogen Activated Protein Kinase) [16]. IFN-активированные JAK регулируют фосфорилирование Vav или других GEF (гуанин-нуклеотидные факторы обмена), что приводит к последующей активации Rac1, который может регулировать путь передачи сигналов p38MAPK. Затем активируется MAPKKK (киназа-киназа MAPK), которая регулирует последующую активацию киназ MAPK MAPKK3 и MAPKK6, которые непосредственно фосфорилируют p38, что приводит к его активации.Активированный p38 впоследствии регулирует активацию нескольких нижестоящих эффекторов, включая MAPKAPK2 (MAPK-Activated Protein Kinase-2), MAPKAPK3, MSK1 / 2 (Mitogen- and Stress-Activated Kinase) и MNK1 (MAPK-Intecting Protein Kinase-1) [17 ]. Специфические факторы транскрипции, которые регулируются p38s, включают CREB (белок, связывающий цАМФ) и гистон-h4. Активированные TYK2 и JAK1 регулируют фосфорилирование тирозина IRS1 и IRS2 (субстрат рецептора инсулина), которые обеспечивают сайты стыковки для доменов Sh3 (SRC Homology-2) регуляторной субъединицы (p85) PI3K (фосфатидилинозитол 3-киназы).PI3K впоследствии активируется и регулирует последующую активацию mTOR (мишень рапамицина млекопитающих), которая опосредует инициацию трансляции мРНК. В свою очередь, mTOR регулирует фосфорилирование (активацию) p70S6K (киназа p70S6), которая затем фосфорилирует RPS6 (рибосомный белок S6), что приводит к инициации трансляции мРНК. mTOR также регулирует фосфорилирование репрессора трансляции 4EBP1 (Eukaryotic Translation-Initiation Factor 4E (EIF4E) -Binding Protein-1). Такое фосфорилирование приводит к его дезактивации и последующей диссоциации от EIF4E, что позволяет инициировать кэп-зависимую трансляцию мРНК.
IFN типа I эффективны при лечении злокачественных новообразований, вирусных инфекций и аутоиммунных заболеваний. IFN-Alpha играет роль в лечении различных новообразований, особенно гематологического происхождения, то есть миелоидной и лимфоидной этиологии [17]. IFN-бета — единственный интерферон, который, как было показано, оказывает положительное воздействие при лечении рассеянного склероза. IFN-Gamma является абсолютным требованием для устойчивости к острой приобретенной инфекции Toxoplasma gondii и развитию TE (токсоплазматического энцефалита) на поздней стадии инфекции [18].Таким образом, этот уникальный набор биологических активностей IFNs вызвал значительный интерес к его потенциалу для лечения вирусных инфекций, рака и иммунологических нарушений. Действительно, введение IFN в настоящее время является методом выбора для лечения некоторых видов рака, особенно лейкозов и саркомы Капоши, ревматоидного артрита и вирусных инфекций, таких как хронический гепатит B и C. Однако многие другие типы рака и вирусные инфекции лишь частично поддаются лечению. IFN-терапия. Молекулярные механизмы, которые определяют противовирусную и антипролиферативную чувствительность или устойчивость к IFN, еще предстоит полностью выяснить.
IRF-1, RIG-I и MDA5 проявляют мощную противовирусную активность против норовируса, координированно индуцируемого различными типами интерферонов
Основные моменты
Abstract
Норовирус представляет собой основную причину острого небактериального гастроэнтерита во всем мире. У пациентов с ослабленным иммунитетом он несет высокий риск вызвать хроническую инфекцию со значительной заболеваемостью и смертностью.Отсутствие специфического лечения побуждает к разработке антиноровирусных агентов. В этом исследовании мы изучили роль ответа интерферона (IFN) и оценили противовирусную активность различных IFN против репликации норовируса человека (HuNoV) с использованием модели репликона HuNoV. Мы обнаружили, что репликация РНК HuNoV была чувствительна ко всем типам IFN, включая IFNα (тип I), IFNγ (тип II), IFNλ1 и 3 (тип III). IFNs канонически индуцируют стимулированные интерфероном гены (ISG) к проявлению их противовирусной активности.Путем профилирования подмножества важных ISG человека с использованием подхода сверхэкспрессии мы определили RTP4 и HPSE как умеренные анти-норовирусные ISG, тогда как IRF-1, RIG-I (также известный как DDX58 ) и MDA5 (также известный как IFIh2 ) были идентифицированы как сильные анти-норовирусные эффекторы. Интересно, что IFN типа I и III координированно индуцировали IRF-1, RIG-I и MDA5; тогда как IFN типа II преимущественно индуцировал IRF-1 для проявления их анти-норовирусной активности. Комбинация различных IFN показала, что IFNγ работает совместно с IFN типа I или типа III, индуцируя ISG и впоследствии подавляя репликацию HuNoV.Следует отметить, что репликация HuNoV не влияла на противовирусный ответ IFN. Таким образом, мы показали сильную анти-норовирусную активность различных типов IFN и определили ключевые анти-норовирусные эффекторы. Эти результаты важны для понимания взаимодействия норовируса с хозяином и разработки противовирусных методов лечения.
Ключевые слова
Норовирус
Интерферон
IRF-1
RIG-I
MDA5
Сокращения
HBGAАнтиген гисто-группы крови
ВИЧГен, стимулированный вирусом иммунодефицита человека
ISG-9000 -1регуляторный фактор интерферона-1
MDA5ген, связанный с дифференцировкой меланомы 5
PRRрецепторов распознавания образов
PBSфосфатно-солевой буфер
RIG-Iиндуцируемый ретиноевой кислотой ген I
Рекомендуемые статьи Цитирующие статьи (0)
Посмотреть аннотацию© 2018 Авторы.Опубликовано Elsevier B.V.
Рекомендуемые статьи
Цитирующие статьи
IFN-λ3 ингибирует ВИЧ-инфекцию макрофагов через путь JAK-STAT
Абстрактные
Фон
Интерферон лямбда 3 (IFN-λ3) представляет собой недавно идентифицированный цитокин с противовирусной активностью, и его однонуклеотидный полиморфизм тесно связан с эффективностью лечения и развитием хронической вирусной инфекции гепатита С. Таким образом, мы исследовали способность IFN-λ3 ингибировать репликацию ВИЧ и возможные механизмы анти-ВИЧ действия IFN-λ3 в макрофагах человека.
Основные результаты
В различных условиях (до, во время и после ВИЧ-инфекции) IFN-λ3 значительно ингибировал репликацию вируса в макрофагах, что было связано с индукцией множества противовирусных клеточных факторов (ISG56, MxA, OAS-1, A3G / F и тетерин). ) и регуляторные факторы IFN (IRF-1, 3, 5, 7 и 9). Это действие IFN-λ3 против ВИЧ может быть нарушено ингибитором JAK-STAT. Кроме того, обработка макрофагов IFN-λ3 индуцировала экспрессию toll-подобного рецептора 3 (TLR3) и двух ключевых адаптеров (MyD88 и TRIF) при активации пути IFN I типа.Однако ВИЧ-инфекция нарушила опосредованную IFN-λ3 индукцию ключевых элементов сигнального пути JAK-STAT.
Выводы
Эти данные показывают, что IFN-λ3 выполняет свою функцию против ВИЧ, активируя в макрофагах опосредованный путем JAK-STAT врожденный иммунитет. В будущем необходимо провести in vivo исследований, чтобы изучить возможность разработки терапии ВИЧ-инфекции на основе IFN-λ3.
Образец цитирования: Лю M-Q, Zhou D-J, Wang X, Zhou W, Ye L, Li J-L, et al.(2012) IFN-λ3 ингибирует ВИЧ-инфекцию макрофагов посредством пути JAK-STAT. PLoS ONE 7 (4): e35902. https://doi.org/10.1371/journal.pone.0035902
Редактор: Джиалин Чарльз Чжэн, Медицинский центр Университета Небраски, Соединенные Штаты Америки
Поступила: 8 сентября 2011 г .; Одобрена: 23 марта 2012 г .; Опубликовано: 27 апреля 2012 г.
Авторские права: © 2012 Liu et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Это исследование было поддержано Национальным институтом здравоохранения / Национальным институтом злоупотребления наркотиками (№ DA12815, DA22177 и DA27550) и Национальным фондом естественных наук Китая (№ 81001303). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Интерфероновые лямбды (IFN-λ) представляют собой класс недавно идентифицированных членов семейства IFN.Подсемейство IFN-λ состоит из трех структурно связанных цитокинов (IFN-λ1, IFN-λ2, IFN-λ3), которые также называются интерлейкином-29 и интерлейкином-28A / B (IL-29, IL-28A, IL-28B ) соответственно [1], [2]. IFN-λ может быть активирован после вирусных инфекций или активации толл-подобных рецепторов (TLR) [3], [4]. Экспрессия IFN-λ может индуцировать противовирусные факторы для подавления репликации широкого спектра вирусов [4]. IFN-λ функционально напоминает IFN типа I, вызывая противовирусную защиту in vitro [1], [2], [5], [6], а также in vivo [7].Однако, в отличие от рецепторов IFN типа I, которые широко экспрессируются в большинстве типов клеток, включая клетки головного мозга, характер экспрессии рецепторов IFN-λ более специфичен для клеток. Таким образом, IFN-λ имеет меньше побочных эффектов, чем IFN-α [8], что считается значительным преимуществом перед IFN-α. IFN-λ связывается с двумя своими рецепторами, которые активируют сигнальный путь JAK-STAT, что приводит к фосфорилированию белков STAT и образованию комплекса ISGF3 [9]. Образованный комплекс ISGF3 связывается с IFN-стимулированным ответным элементом (ISRE) и вызывает ответы хозяина на вирусные инфекции [10].Потенциальная клиническая важность IFN-λ как нового противовирусного терапевтического агента недавно стала очевидной. Несколько групп [11] — [14] сообщили, что эндогенная система IFN-λ связана с вызванным лечением клиренсом вируса гепатита C (HCV). Это открытие еще больше повысило интерес к изучению противовирусных механизмов IFN-λ, особенно IFN-λ3.
Хотя сообщалось, что два члена семейства IFN-λ, IFN-λ1 и IFN-λ2, могут ингибировать репликацию ВИЧ в макрофагах [6], тем не менее неясно, обладает ли IFN-λ3 функцией против ВИЧ как хорошо.В этом исследовании мы изучили, обладает ли IFN-λ3 противовирусной способностью против ВИЧ-инфекции макрофагов. Мы также исследовали механизмы, участвующие в опосредованных IFN-λ3 действиях против ВИЧ.
Материалы и методы
Клетки и вирусы
Образцы периферической крови были получены от здоровых доноров и протестированы на антитела к ВИЧ. Институциональный наблюдательный совет Университета Темпл одобрил это исследование, и от всех испытуемых было получено письменное информированное согласие.Все расследования проводились в соответствии с принципами, изложенными в Хельсинкской декларации. РВМС выделяли градиентом фиколла и дополнительно инкубировали с модифицированной Дульбекко средой Игла (DMEM) в покрытых желатином колбах в течение 45 мин при 37 ° C для сбора моноцитов. После отделения 10 мМ ЭДТА моноциты промывали DMEM и ресуспендировали в 10% DMEM, включая 10% фетальную бычью сыворотку, глутамин (2 ммоль / мл), пенициллин (100 Ед / мл), стрептомицин (100 мкг / мл) и заменимые аминокислоты.Очистку моноцитов определяли неспецифическим окрашиванием эстеразой и анализом сортировки клеток с активацией флуоресценции с использованием моноклональных антител против CD14 (Leu-M3) и липопротеинов низкой плотности, специфичных для моноцитов и макрофагов [6]. Моноциты высевали в 96-луночный планшет (10 5 клеток / лунку) или 48-луночный планшет (2,5 × 10 5 клеток / лунку) и дифференцировали в макрофаги после культивирования в течение 7 дней. Штамм ВИЧ Bal (R5) был получен в рамках Программы исследований и референс-реагентов по СПИДу Национального института здравоохранения (Бетесда, Мэриленд).
Реактивы
Рекомбинантные человеческие IFN-λ1 и IFN-λ2 были приобретены у PeproTech Inc. (Rocky Hill, NJ). Рекомбинантный человеческий IFN-λ3 был приобретен в R&D Systems, Inc. (Миннеаполис, Миннесота), чистота IFN-λ3 аналогична чистоте IFN-λ1 и IFN-λ2. Ингибитор I JAK был приобретен у EMD Chemicals, Inc. (Гиббстаун, Нью-Джерси) и растворен в 1% диметилсульфоксиде (ДМСО). Моноклональное антитело к ВИЧ p24 было получено от NIH AIDA Research & Reference Reagent Program.
ВИЧ-инфекция и лечение IFN-λ
Семидневные культивированные макрофаги инкубировали с IFN-λ либо за 24 часа до инфицирования ВИЧ Bal, либо одновременно с ВИЧ-инфекцией, либо через 4 дня после инфицирования ВИЧ.Продуктивную ВИЧ-инфекцию определяли на 12-й день после инфицирования. В экспериментах, в которых изучали влияние дозы и времени IFN-λ3, IFN-λ3 добавляли к культурам через 4 дня после инфицирования и постоянно присутствовали в культурах. Чтобы определить роль пути JAK-STAT в действии IFN-λ3 против ВИЧ, ингибитор JAK I был добавлен в культуру клеток за 1 час до обработки IFN-λ3.
Анализ активности обратной транскриптазы (RT)
АктивностьОТ ВИЧ определяли по методике Willey et al.[15] с некоторыми изменениями [6], [16]. Вкратце, 10 мкл супернатанта, собранного из культур ВИЧ-инфицированных макрофагов, добавляли к 50 мкл коктейля, содержащего поли (A), олиго (dT), MgCl 2 , Nonidet P-40 и [ 32 P] dTTP и инкубировали в течение ночи при 37 ° C. Тридцать микролитров реакционной смеси наносили на бумагу DE 81 и сушили на воздухе. Затем фильтры промывали четыре раза свежим стандартным цитратом физиологического раствора (SSC, содержащий 0,3 моль / л NaCl, 0,03 моль / л цитрата натрия, PH 7) и 100% этанолом.После сушки фильтры помещали в сцинтилляционные флаконы и измеряли радиоактивность с помощью жидкостного сцинтилляционного анализатора (PerkinElmer Tri-Carb 2810TR, Downers Grove, IL60515, США).
Экстракция РНК и ОТ-ПЦР в реальном времени
Общий клеточный нуклеотид экстрагировали из макрофагов с использованием реагента TRI® (SIGMA-ALDRICH, Inc. St. Louis, MO 63178, США), как описано ранее [6]. Тотальную РНК (1 мкг) подвергали обратной транскрипции с использованием системы RT (Promega, Madison, WI) в течение 1 ч при 42 ° C.Полученную кДНК использовали в качестве матрицы для количественной оценки ПЦР в реальном времени. Используя супермикс iQ SYBR Green (Bio-Rad Laboratories, Геркулес, Калифорния), информационные РНК (мРНК) в макрофагах были обнаружены для противовирусных факторов (интерферон-стимулированный ген 56 (ISG-56), устойчивость к миксовирусу-1 (MxA), обструктивная апноэ сна-1 (OAS-1), APOBEC3G / APOBEC3F (A3G / F), тетерин, регуляторный фактор IFN (IRF) -1, IRF-3, IRF-5, IRF-7, IRF-9, толл-подобный рецептор (TLR) -3, TLR-7, TLR-9, рецепторы IFN-λ3 (IL-28Rα, IL-10Rβ), цитокины сигнального пути IFN-λ3 (ген 88 первичного ответа миелоидной дифференцировки (MyD88), TIR-домен, содержащий адаптер-индуцирующий интерферон-β (TRIF), преобразователь сигнала и активатор транскрипции (STAT-1/2)) и глицеральдегид-3-фосфатдегидрогеназа (GAPDH).Праймеры, использованные для вышеуказанных параметров, были перечислены в таблице 1. Уровни мРНК GAPDH использовали в качестве эндогенного эталона для нормализации количеств мРНК-мишени и представляли как изменение индукции по сравнению с необработанными контрольными клетками.
Непрямой иммунофлуоресцентный анализ на ВИЧ p24
ВИЧ-инфицированных Bal макрофагов, обработанных IFN-λ3 или без него, промывали фосфатно-солевым буфером (PBS) и фиксировали в 4% параформальдегиде (EM Science) в течение 20 минут при комнатной температуре.Затем клетки пермеабилизировали 0,2% тритоном в течение 10 минут и блокировали в блокирующем растворе (10% фетальная телячья сыворотка, 1% пенициллин / стрептомилин и 0,05% азид натрия в MEM) в течение 1 часа. Антиген р24 ВИЧ детектировали с использованием мышиных моноклональных антител против ВИЧ-р24 и козьих антител против мыши, конъюгированных с Alexa Fluor 488 (Invitrogen). Ядра клеток контрастировали красителем Hoechst. Микроскопический анализ выполняли с помощью флуоресцентного микроскопа (Olympus).
Статистический анализ
При необходимости данные были выражены как средние значения ± стандартное отклонение (SD) по крайней мере для трех повторностей образцов.Анализ данных проводился с использованием двустороннего t-критерия Стьюдента. Статистический анализ выполняли с помощью статистического программного обеспечения Graphpad Instat (Graphpad Software Inc., Сан-Диего, Калифорния), и статистическая значимость была определена как P <0,05 .
Результаты
IFN-λ3 ингибирует репликацию ВИЧ в макрофагах
Сначала мы исследовали влияние IFN-λ3 на репликацию ВИЧ в макрофагах. При добавлении к ВИЧ-инфицированным культурам макрофагов IFN-λ3 значительно ингибировал активность ОТ ВИЧ в дозе (рис.1А) и в зависимости от времени (рис. 1В). Этот ингибирующий эффект IFN-λ3 на ВИЧ наблюдался при трех различных условиях лечения (фиг. 1C). Наибольшее ингибирование ВИЧ наблюдалось в культурах макрофагов, предварительно обработанных IFN-λ3 (до заражения ВИЧ) (рис. 1С). Для поддержания ИФН-λ3 в культурах ВИЧ-инфицированных клеток было необходимо, так как изъятие IFN-λ3 из инфицированных культур приводило к отскоку вируса (рис. 1С). Ингибирование ВИЧ IFN-λ3 было также подтверждено непрямым иммунофлюоресцентным окрашиванием антителами против антигена ВИЧ p24 (рис.2).
Рисунок 1. Влияние IFN-λ3 на ВИЧ-инфекцию макрофагов человека.
A. Дозозависимый эффект IFN-λ3 на ВИЧ-инфекцию. Семидневные культивированные макрофаги инфицировали штаммом ВИЧ Bal в течение 4 дней, а затем обрабатывали IFN-λ3 в указанных концентрациях. Активность ОТ ВИЧ в супернатанте анализировали на 12 день после инфицирования. B. Зависящий от времени эффект IFN-λ3 на ВИЧ-инфекцию. Семидневные культивированные макрофаги инфицировали штаммом ВИЧ Bal в течение 4 дней, а затем обрабатывали IFN-λ3 в концентрации 100 нг / мл.Исследовали активность ОТ ВИЧ в супернатанте в указанный момент времени. C. Влияние IFN-λ3 на ВИЧ-инфекцию и репликацию в четырех различных условиях. Макрофаги предварительно обрабатывали IFN-λ3 (100 нг / мл) в течение 24 часов, а затем инфицировали штаммом ВИЧ Bal (до) или совместно инкубировали с IFN-λ3 (100 нг / мл) и штаммом ВИЧ Bal при в то же время (во время) или через 4 дня после ВИЧ-инфекции макрофаги в указанных выше условиях обрабатывали IFN-λ3 (100 нг / мл) с отменой IFN-λ3 или без нее.Уровни активности ОТ ВИЧ в супернатанте анализировали на 12 день после инфицирования. Представленные данные представляют собой среднее значение ± стандартное отклонение для трех повторностей культуры и выражены как% от контроля (без обработки IFN-λ3, среднее значение которого определено как 100) ( * P <0,05, ** P <0,01 ) .
https://doi.org/10.1371/journal.pone.0035902.g001
Рис. 2. Непрямой иммунофлуоресцентный анализ на p24 ВИЧ.
Семидневные культивированные макрофаги инфицировали штаммом ВИЧ Bal или без него в течение 2 часов и культивировали в течение 12 дней в присутствии или в отсутствие IFN-λ3 (100 нг / мл).Затем клетки промывали и фиксировали 4% параформальдегидным фиксатором и обрабатывали 0,2% тритоном. ВИЧ p24 в инфицированных макрофагах окрашивали антисывороткой к ВИЧ p24 (зеленый цвет), а ядра клеток окрашивали Hoechst и наблюдали с помощью флуоресцентного микроскопа. (NC, отрицательный контроль; ПК, положительный контроль).
https://doi.org/10.1371/journal.pone.0035902.g002
IFN-λ3 индуцирует ключевые элементы пути JAK-STAT
Чтобы определить механизм опосредованной IFN-λ3 активности против ВИЧ в макрофагах, мы исследовали влияние IFN-λ3 на экспрессию TLR, а также несколько ключевых элементов в сигнальном пути IFN-λ как в неинфицированных, так и в инфицированных макрофагах.Сначала мы определили, модулирует ли обработка IFN-λ3 экспрессию TLR (TLR3, 7, 9) в макрофагах. Обработка неинфицированных макрофагов IFN-λ3 может индуцировать экспрессию TLR7 и TLR9 (фиг. 3A). Более того, IFN-λ3 индуцировал экспрессию TLR3 в неинфицированных макрофагах в 40 раз (фиг. 3A). Затем мы исследовали, регулирует ли IFN-λ3 экспрессию регуляторных факторов IFN (IRF). Как показано на фиг. 3B, IFN-λ3 индуцировал экспрессию нескольких ключевых IRF, в частности IRF7 в неинфицированных макрофагах (фиг.3Б). Поскольку IFN-λ посредством связывания с IL-28Rα и IL-10Rβ активирует путь JAK-STAT [17], а активация TLR вызывает экспрессию IFN типа I [18], мы дополнительно исследовали влияние IFN-λ3 на IFN типа I. сигнальный путь. Как показано на фиг. 3C, экспрессия IL-28Rα и STAT-1/2 в значительной степени индуцировалась IFN-λ3 в неинфицированных макрофагах. Кроме того, обработка неинфицированных макрофагов IFN-λ3 усиливала экспрессию двух ключевых адаптеров (MyD88 и TRIF) при активации пути IFN I типа (рис.3С). Поскольку ВИЧ-инфекция / репликация может иметь отрицательный эффект на экспрессию противовирусных клеточных факторов, мы исследовали влияние ВИЧ на IFN-λ3-опосредованную индукцию ключевых элементов пути JAK-STAT в инфицированных макрофагах. Как показано на фиг. 4, хотя IFN-λ может индуцировать экспрессию TLR, а также других противовирусных факторов в инфицированных макрофагах, степень индукции была ниже (фиг. 4), чем в неинфицированных клетках (фиг. 3).
Рисунок 3. Влияние IFN-λ3 на ключевые элементы пути JAK-STAT в неинфицированных макрофагах.
Семидневные культивированные макрофаги обрабатывали IFN-λ3 (100 нг / мл) или без него в течение 24 часов. Суммарные нуклеотиды экстрагировали из макрофагов и подвергали анализу полимеразной цепной реакции с обратной транскрипцией (RT-PCR) в реальном времени для определения экспрессии мРНК TLR ( A ), IRF ( B ) и ключевых элементов передачи сигналов IFN-λ3. путь ( C ). Представленные данные представляют собой среднее значение ± стандартное отклонение для трех повторностей культуры ( * P <0,05, ** P <0,01 ).
https: // doi.org / 10.1371 / journal.pone.0035902.g003
Рис. 4. Влияние IFN-λ3 на ключевые элементы пути JAK-STAT в ВИЧ-инфицированных макрофагах.
Семидневные культивированные макрофаги инфицировали штаммом ВИЧ Bal в течение 2 часов. IFN-λ3 (100 нг / мл) добавляли к культурам через 4 дня после инфицирования в течение 24 часов. Суммарные нуклеотиды экстрагировали из макрофагов и подвергали анализу полимеразной цепной реакции с обратной транскрипцией (RT-PCR) в реальном времени для определения экспрессии мРНК TLR ( A ), IRF ( B ) и ключевых элементов передачи сигналов IFN-λ3. путь ( C ).Семидневные культивированные макрофаги, не инфицированные штаммом ВИЧ Bal и не обработанные IFN-λ3, были выбраны в качестве контроля для справедливого сравнения с данными на фиг.3. Показанные данные представляют собой среднее значение ± стандартное отклонение для трех повторностей культивирования ( * P <0,05. , ** P <0,01 ).
https://doi.org/10.1371/journal.pone.0035902.g004
IFN-λ3 более эффективно подавляет ВИЧ, чем IFN-λ1 и IFN-λ2
Было показано, чтоIFN-λ1 или IFN-λ2 ингибируют репликацию ВИЧ в макрофагах [6], поэтому мы сравнили анти-ВИЧ способность IFN-λ3 с IFN-λ1 или IFN-λ2.Как показано на фиг. 5A, ингибирующий эффект IFN-λ3 на ВИЧ в макрофагах более мощный, чем IFN-λ1 или IFN-λ2. Кроме того, IFN-λ3 был наиболее мощным индуктором клеточных противовирусных факторов (ISG-56, MxA, OAS-1, A3G / F и тетерин) среди трех членов семейства IFN-λ (фиг. 5B).
Рис. 5. Влияние IFN-λ1, IFN-λ2 и IFN-λ3 на ВИЧ.
A. Семидневные культивированные макрофаги инфицировали штаммом ВИЧ Bal в течение 4 дней, а затем обрабатывали IFN-λ1 / 2 (100 нг / мл) или IFN-λ3 (100 нг / мл) или без них в течение 8 дней. дней.Активность ОТ ВИЧ в супернатанте анализировали на 12 день после инфицирования. Данные были выражены как относительные уровни ОТ ВИЧ (%) к контролю (без обработки IFN-λ, который определен как 100). B. Семидневные культивированные макрофаги обрабатывали IFN-λ1 / 2 (100 нг / мл) или IFN-λ3 (100 нг / мл) или без них в течение 24 часов, и общий нуклеотид макрофагов экстрагировали для реального анализа. время ОТ-ПЦР-анализ экспрессии мРНК ISG-56, MxA, OAS-1, A3G / F, тетерина и GAPDH. Данные были выражены в виде уровней мРНК для факторов анти-ВИЧ относительно (кратных) к контролю (без обработки IFN-λ, который определяется как 1).( * P <0,05, ** P <0,01 ).
https://doi.org/10.1371/journal.pone.0035902.g005
Активность IFN-λ3 против ВИЧ зависит от пути JAK-STAT
Чтобы изучить механизмы активности IFN-λ3 против ВИЧ, мы исследовали роль пути JAK-STAT в действии IFN-λ3 против ВИЧ. Как показано на фиг. 6A, было обнаружено, что активность IFN-λ3 против ВИЧ значительно подавляется ингибитором (ингибитор I JAK) пути JAK-STAT.Кроме того, лечение ингибитором JAK I нарушило индуцирующие эффекты IFN-λ3 на несколько ключевых противовирусных клеточных факторов, включая ISG-56, MxA, OAS-1, A3G / F и тетерин (фиг. 6B).
Рис. 6. Влияние ингибитора JAK-STAT на действие IFN-λ3.
A. Влияние ингибитора JAK-STAT на действие IFN-λ3 против ВИЧ. Макрофаги инфицировали штаммом ВИЧ Bal в течение 4 дней, а затем инкубировали с ингибитором JAK I (1 мкМ) в течение 1 часа перед обработкой IFN-λ3 (100 нг / мл).Активность ОТ ВИЧ в супернатанте измеряли на 12 день после инфицирования. B. Влияние ингибитора JAK-STAT на IFN-λ3-опосредованную индукцию клеточных противовирусных факторов. Макрофаги инкубировали с ингибитором JAK (1 мкМ) в течение 1 часа перед обработкой IFN-λ3 (100 нг / мл) в течение 24 часов. Экспрессия ISG-56, MxA, OAS-1, A3G / F и тетерина была указана как увеличение индукции (кратность) по сравнению с клетками, не обработанными IFN-λ3, что нормализовано к уровням GAPDH. Значения были выражены как среднее значение ± стандартное отклонение для трех повторностей культуры ( * P <0.05, ** P <0,01 ).
https://doi.org/10.1371/journal.pone.0035902.g006
Обсуждение
По сравнению с IFN-λ1 или IFN-λ2, IFN-λ3 считается более важным для терапии против HCV, поскольку его генетическая вариация трех однонуклеотидных полиморфизмов (SNP) (rs12979860, rs12980275 и rs8099917) в восходящем направлении от гена IFN-λ3 (IL-28B) тесно связан с результатами лечения Peg-IFNα-2a в сочетании с терапией рибавирином [11] — [14]. Хотя противовирусная функция IFN-λ1 или IFN-λ2 была продемонстрирована [6], еще предстоит определить, обладает ли IFN-λ3 способностью подавлять ВИЧ.В настоящем исследовании мы предоставили экспериментальные доказательства того, что, подобно IFN-λ1 и IFN-λ2, лечение IFN-λ3 приводило к ингибированию ВИЧ-инфекции макрофагов. Когда макрофаги были предварительно обработаны один раз IFN-λ3, наблюдалось почти полное ингибирование ВИЧ макрофагов (рис. 1). Присутствие IFN-λ3 в культурах необходимо для поддержания подавления ВИЧ, поскольку вывод IFN-λ3 из культур привел к возобновлению репликации вируса (рис. 1C). Этот ингибирующий эффект лечения IFN-λ3 наблюдался даже после того, как ВИЧ-инфекция была инициирована в макрофагах (рис.1). Мы также продемонстрировали, что IFN-λ3 оказался более эффективным в ингибировании репликации ВИЧ в макрофагах, чем IFN-λ1 или IFN-λ2 (рис. 5). Эта повышенная способность ингибировать ВИЧ может быть связана с его более сильным воздействием на несколько ключевых клеточных противовирусных факторов в макрофагах, чем IFN-λ1 или IFN-λ2 (рис. 5B).
IFN-λ функционально напоминает IFN I типа по противовирусной защите [19]. Несмотря на перекрывающиеся противовирусные функции IFN-λ с IFN типа I, более строгий паттерн экспрессии рецептора IFN-λ (IL-28Rα) предполагает, что IFN-λ играет более специализированную роль в противовирусной защите хозяина против вирусов в подмножестве клетки [20], [21].Кроме того, механизмы, с помощью которых IFN-λ устанавливает противовирусное состояние, не так хорошо охарактеризованы, как механизмы для IFN-α / β. Наши исследования показали, что IFN-λ3 посредством нескольких механизмов подавляет ВИЧ-инфекцию и репликацию. Сначала мы исследовали, активирует ли IFN-λ3 сигнальный путь JAK-STAT, вызывая антивирусное состояние в макрофагах. Обработка макрофагов IFN-λ3 усиливала экспрессию либо ключевых факторов пути JAK-STAT, таких как STAT1 и STAT2 (рис. 3C, 4C), либо известных ISG, расположенных ниже пути JAK-SATA, включая ISG-56, MxA и OAS-1 (рис.5Б). Роль пути JAK-STAT в активности IFN-λ3 против ВИЧ была дополнительно подтверждена наблюдением, что блокирование сигнального пути JAK-STAT ингибитором JAK может поставить под угрозу действие IFN-λ3 на ВИЧ и значительно снизить экспрессию IFN-λ3-индуцированных противовирусных факторов (рис. 6). Мы также продемонстрировали, что APOBEC3G и APOBEC3F, специфические клеточные факторы против ВИЧ, индуцировались IFN-λ через путь JAK-STAT (фиг. 5B и 6B). Кроме того, тетерин, трансмембранный белок, который специфически ингибирует ВИЧ-инфекцию, предотвращая его высвобождение из инфицированных клеток, также индуцировался IFN-λ через путь JAK-STAT (рис.5Б и 6Б). Таким образом, путь JAK-STAT играет решающую роль в анти-ВИЧ-активности IFN-λ3 в макрофагах, что согласуется с предыдущими выводами, показывающими, что IFN-λ3 через активацию пути JAK-STAT оказывает свое антивирусное действие. -HCV функция в гепатоцитах человека [10].
TLRимеют решающее значение для врожденных иммунных ответов на патогены, поскольку они распознают и отвечают на PAMP, что приводит к активации внутриклеточных сигнальных путей. Среди одиннадцати идентифицированных TLR человека три TLR (TLR-3, 7 и 9) играют ключевую роль в опосредованном вирусами врожденном иммунитете, поскольку они специфически распознают вирусную РНК или ДНК и инициируют пути передачи противовирусных сигналов в макрофагах [22], [22], [22] 23].Наши исследования показали, что IFN-λ3 может значительно индуцировать экспрессию TLR3, а также связанных с TLR3 адаптеров, таких как MyD88 и TRIF (рис. 3A, 3C, 4A и 4C). Эти наблюдения обеспечивают дополнительный механизм опосредованного IFN-λ3 действия против ВИЧ, поскольку передача сигналов TLR3 играет решающую роль в раннем врожденном иммунном ответе на вирусные инфекции, включая ВИЧ [24].
Наше дальнейшее исследование показало, что IFN-λ3 индуцировал экспрессию IRF (IRF-1, 3, 5, 7 и 9) в макрофагах, инфицированных или не инфицированных ВИЧ (рис.3Б и 4Б). IRF являются важными регуляторами противовирусного иммунитета, опосредованного IFN I и III типов. Эти регуляторы не только распознают элементы промотора ДНК для модуляции генов IFN I и III типов, но также регулируют IFN-стимулированный ответный элемент (ISRE) в некоторых ISG, что приводит к индукции антивирусного состояния [25], [26] . Недавние исследования [27] — [29] дополнительно продемонстрировали важную роль IRF-3, IRF-5 и IRF-7 в активации экспрессии IFN типа I и III, что имеет решающее значение для производства последующих противовирусных препаратов. такие факторы, как ISG-56, MxA, OAS-1, tetherin (рис.6).
Взятые вместе, наши данные предоставили убедительные доказательства того, что IFN-λ3 через активацию путей TLR3 и JAK-STAT ингибирует ВИЧ-инфекцию макрофагов. Хотя точные клеточные и молекулярные механизмы, с помощью которых IFN-λ3 ингибирует репликацию ВИЧ, еще предстоит определить, индукция множественных клеточных факторов рестрикции против ВИЧ должна составлять большую часть опосредованной IFN-λ3 активности против ВИЧ. Эти анти-ВИЧ механизмы действия IFN-λ3 предлагают привлекательную альтернативу лечению ВИЧ, поскольку для ВИЧ было бы чрезвычайно трудно выработать устойчивость к действиям IFN-λ3, которые подавляют вирус на различных этапах его репликации в контексте хозяина. клеточный врожденный иммунитет.Однако необходимы дальнейшие исследования, чтобы определить влияние IFN-λ3 на ВИЧ в системах ex vivo и in vivo . Эти дополнительные исследования должны изучить возможность разработки терапии на основе IFN-λ3 для людей, инфицированных ВИЧ.
Вклад авторов
Задумал и спроектировал эксперименты: MQL DJZ XW WZ LY JLL YZW WZH. Проведены эксперименты: MQL XW LY JLL. Проанализированы данные: MQL XW. Предоставленные реагенты / материалы / инструменты анализа: MQL XW LY.Написал статью: MQL DJZ XW WZH.