Регулирующая функция налогов и ее характеристика.
Сущность и внутреннее содержание налогов проявляется в функциях, которые они выполняют. Регулирующая функция позволяет государству активно воздействовать на экономические и социальные процессы. Она связана с распределением налоговых платежей между юридическими и физическими лицами, отраслями экономики. Данная функция позволяет регулировать доходы разных групп населения. Налоговое регулирование реализуется через систему льгот и систему налоговых платежей.
Цель
применения налоговых льгот- сокращение
размера налоговых обязательств
плательщика. Льготы делятся на изъятия,
скидки, налоговый кредит. Регулирующая
функция направлена на регулирование
финансово- хозяйственной деятельности
производителей товаров через систему
налоговых платежей для восстановления
израсходованных ресурсов. Эти отличия
имеют четкую отраслевую направленность-
налог на пользование объектами животного
мира, водный налог, земельный налог.
+Регулирующая функция проявляется не только в сфере производства, но и через платежеспособность физических лиц- на рынке спроса и предложения на товары.
Контрольная функция налогов и ее характеристика.
Сущность
и внутреннее содержание налогов
проявляется в функциях, которые они
выполняют. Контрольная функция заключается
в том, что налоги выступают своеобразным
зеркалом экономических процессов. Она
состоит в том, что появляется возможность
отражения налоговых поступлений.
Благодаря этой функции оценивается
эффективность налогового механизма,
обеспечивается контроль за движением
финансовых ресурсов. Контрольная функция
зависит от налоговой дисциплины. Смысл
в том, чтобы налогоплательщики своевременно
и в полном объеме уплачивали установленные
законодательством налоги. Анализ
динамики налоговых поступлений позволяет
составить картину о процессах, происходящих
в экономике, о влиянии налоговой системы
на национальное хозяйство, о достаточности
средств для финансирования бюджетных
мероприятий.
+Контрольная функция реализуется органами, осуществляющими контроль за правильным исчислением, уплатой налогов- Минфин и таможенные органы.
Особенность формирования налоговой системы Республики Беларусь.
Налоговая система- это совокупность налогов, принципов, форм и методов их установления, взимания, изменения, а также налоговых органов, форм налогового контроля.
Налоговая система РБ- это двухуровневая система, т.е. налоги поступают в республиканский и местный бюджеты. При взимании используются следующие методы: кадастровый; декларационный; административный.
Современная налоговая система РБ характеризуется следующими положительными чертами:
— создана единая правовая для всех субъектов хозяйствования
— применяются общепризнанный набор налоговых платежей
— применяются спец режимы налогообложения для резидентов СЭЗ
+Вместе
с тем, в налоговой системе РБ присущи
следующие недостатки: высокий уровень
налогообложения субъектов хозяйствования;
несвоевременность выхода актов налогового
законодательства; невысокая роль местных
сборов и налогов.
Карта сайта
Главная Обучение Библиотека Карта сайта
|
 2.014.01
2.014.01Что такое НДС?
Налог на добавленную стоимость, или НДС, в Европейском союзе является общим налогом на потребление на широкой основе, взимаемым с добавленной стоимости товаров и услуг. Это применимо более или менее ко всем товарам и услугам, которые покупаются и продаются для использования или потребления в Европейском Союзе. Таким образом, товары, которые продаются на экспорт, или услуги, которые продаются клиентам за границей, обычно не облагаются НДС. И наоборот, импорт облагается налогом, чтобы система оставалась справедливой для производителей ЕС, чтобы они могли на равных конкурировать на европейском рынке с поставщиками, расположенными за пределами Союза.
Это применимо более или менее ко всем товарам и услугам, которые покупаются и продаются для использования или потребления в Европейском Союзе. Таким образом, товары, которые продаются на экспорт, или услуги, которые продаются клиентам за границей, обычно не облагаются НДС. И наоборот, импорт облагается налогом, чтобы система оставалась справедливой для производителей ЕС, чтобы они могли на равных конкурировать на европейском рынке с поставщиками, расположенными за пределами Союза.
Налог на добавленную стоимость
- является общим налогом , который в принципе применяется ко всей коммерческой деятельности, связанной с производством и распределением товаров и предоставлением услуг. Однако, если годовой оборот этого лица меньше определенного предела (порога), который различается в зависимости от государства-члена, лицо не обязано начислять НДС на свои продажи.
- a налог на потребление , потому что в конечном итоге его несет конечный потребитель.
 Это не плата за бизнес.
Это не плата за бизнес. - взимается в процентах от цены, что означает, что фактическое налоговое бремя видно на каждом этапе цепочки производства и распределения.
- собрано дробно , посредством системы частичных платежей, при которой налогообязанные лица (т. е. предприятия, зарегистрированные для уплаты НДС) вычитают из собранного ими НДС сумму налога, которую они уплатили другим налогообязанным лицам при покупках для своей коммерческой деятельности. Этот механизм гарантирует, что налог будет нейтральным независимо от того, сколько транзакций задействовано.
- уплачивается налоговым органам продавцом товаров, который является «налогоплательщиком», но фактически уплачивается покупателем продавцу как часть цены. Таким образом, это косвенный налог.
 Это не плата за бизнес.
Это не плата за бизнес.Почему все страны ЕС используют НДС?
- В то время, когда было создано Европейское Сообщество, первые шесть стран ЕС использовали различные формы косвенного налогообложения, большинство из которых были каскадными налогами. Это были многоступенчатых налогов, каждый из которых взимался с фактической стоимости продукции на каждой стадии производственного процесса, что делало невозможным определение реальной суммы налога, фактически включенной в конечную цену конкретного продукта. Как следствие, всегда существовал риск того, что страны ЕС преднамеренно или случайно будут субсидировать свой экспорт, завышая налоги, возмещаемые при экспорте.
- Было очевидно, что для того, чтобы когда-либо существовал эффективный единый рынок в Европе, требовалась нейтральная и прозрачная система налога с оборота , которая обеспечивала налоговую нейтральность и позволяла возвращать точную сумму налога в момент экспорт. Как поясняется в разделе НДС на импорт и экспорт, НДС позволяет быть уверенным в том, что экспорт полностью и прозрачно не облагается налогом.
 Это были многоступенчатых налогов, каждый из которых взимался с фактической стоимости продукции на каждой стадии производственного процесса, что делало невозможным определение реальной суммы налога, фактически включенной в конечную цену конкретного продукта. Как следствие, всегда существовал риск того, что страны ЕС преднамеренно или случайно будут субсидировать свой экспорт, завышая налоги, возмещаемые при экспорте.
Это были многоступенчатых налогов, каждый из которых взимался с фактической стоимости продукции на каждой стадии производственного процесса, что делало невозможным определение реальной суммы налога, фактически включенной в конечную цену конкретного продукта. Как следствие, всегда существовал риск того, что страны ЕС преднамеренно или случайно будут субсидировать свой экспорт, завышая налоги, возмещаемые при экспорте.Как взимается плата?
НДС, подлежащий уплате при любой продаже, представляет собой процент от продажной цены, но из этой суммы налогоплательщик имеет право вычесть весь налог, уже уплаченный на предыдущем этапе. Таким образом, избегается двойное налогообложение, и налог уплачивается только с добавленной стоимости на каждом этапе производства и распределения. Таким образом, поскольку окончательная цена продукта равна сумме добавленной стоимости на каждом предыдущем этапе, окончательный уплаченный НДС складывается из суммы НДС, уплаченного на каждом этапе.
Таким образом, избегается двойное налогообложение, и налог уплачивается только с добавленной стоимости на каждом этапе производства и распределения. Таким образом, поскольку окончательная цена продукта равна сумме добавленной стоимости на каждом предыдущем этапе, окончательный уплаченный НДС складывается из суммы НДС, уплаченного на каждом этапе.
Зарегистрированным торговцам НДС присваивается номер, и они должны указывать НДС, взимаемый с клиентов в счетах-фактурах. Таким образом, клиент, если он является зарегистрированным торговцем, знает, сколько он может вычесть в свою очередь, а потребитель знает, сколько налога он заплатил за конечный продукт. Таким образом, правильный НДС уплачивается поэтапно, и в определенной степени система является самоконтролируемой.
Пример Стадия 1
Стадия 2
180 евро (оплачивает плавильщик) + 160 евро (оплачивает шахте) + 40 евро (уплачено поставщиком шахте) + 20 евро (уплачено поставщиком плавильному заводу) = 400 евро или правильная сумма НДС при продаже на сумму 2000 евро. |
 Он должен заплатить в казну 200 евро, но, поскольку за тот же отчетный период он купил инструментов на 240 евро, включая НДС на 40 евро, от него требуется заплатить только 160 евро (200 евро минус 40 евро) в казну. Казначейство также получает 40 евро и теперь получает 160 евро, что составляет 200 евро, что является правильной суммой НДС, причитающегося с продажи железной руды.
Он должен заплатить в казну 200 евро, но, поскольку за тот же отчетный период он купил инструментов на 240 евро, включая НДС на 40 евро, от него требуется заплатить только 160 евро (200 евро минус 40 евро) в казну. Казначейство также получает 40 евро и теперь получает 160 евро, что составляет 200 евро, что является правильной суммой НДС, причитающегося с продажи железной руды.
Ставки НДС
Законодательство ЕС требует только, чтобы стандартная ставка НДС была не менее 15%, а сниженная ставка не менее 5% (только для поставок товаров и услуг, указанных в исчерпывающем списке).
Фактические применяемые ставки варьируются в зависимости от стран ЕС и для определенных типов продуктов. Кроме того, некоторые страны ЕС сохранили другие ставки для определенных продуктов.
Наиболее надежным источником информации о текущих ставках НДС для определенного продукта в конкретной стране ЕС является орган по НДС этой страны. Обзор различных ставок, применяемых во всех странах ЕС, представлен в информационном документе ЕС.
Подробнее о ставках НДС
Какова роль Комиссии в применении системы НДС ЕС?
Комиссия несет ответственность за обеспечение правильного применения Директивы по НДС. Каждое государство-член несет ответственность за перенос этих положений в национальное законодательство и их правильное применение на своей территории. Роль Комиссии как «Стража договоров» заключается в обеспечении соответствия национального законодательства и общей практики законодательству ЕС.
Как страны ЕС применяют НДС?
Страны ЕС внедряют общие правила, установленные Директивой по НДС, в свое национальное законодательство. Таким образом, практическое применение и административная практика каждой страны ЕС различаются.
Подробнее о национальных правилах НДС
Может ли Комиссия вмешиваться в конкретные случаи применения Директивы по НДС?
Европейская комиссия не обладает компетенцией решать проблемы отдельных налогоплательщиков в их конкретных случаях, а также давать заключения по фактам.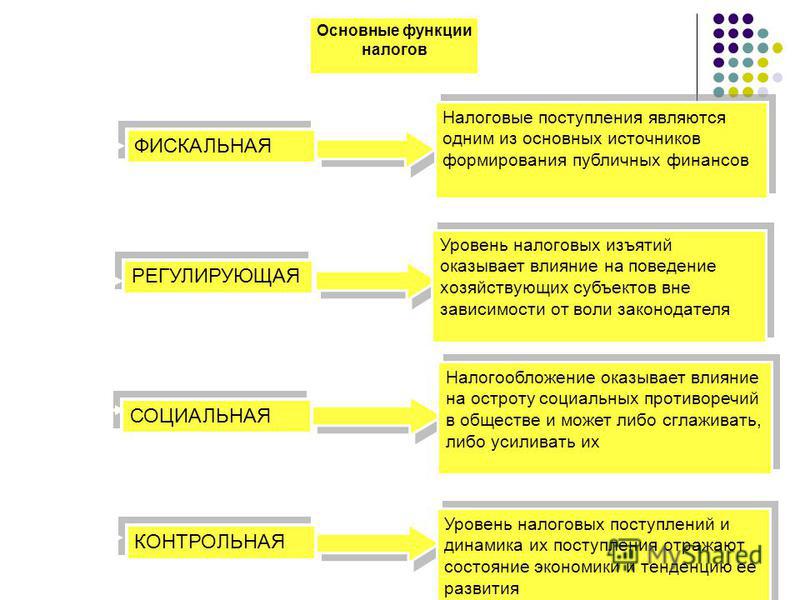
Комиссия может инициировать процедуру нарушения против соответствующего государства-члена. Однако сторонами в этой процедуре считаются только Комиссия и государство-член, за исключением конкретного налогоплательщика. Исход такой процедуры не имеет прямого влияния на конкретные дела.
Таким образом, единственный способ добиться возмещения ущерба в конкретных случаях – это прибегнуть к национальным средствам возмещения ущерба – административным или судебным. Вы также можете отправить свое дело в SOLVIT.
Подробнее о жалобах
См. подробную информацию о правилах НДС по теме
Иммунный гомеостаз висцеральной жировой ткани регулируется перекрестными помехами между адипоцитами и субпопуляциями дендритных клеток Передача сигналов PPARγ и метаболизм: хорошее, плохое и будущее. Нац. Мед. 2013;19: 557–566. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Arokiasamy S., Zakian C., Dilliway J., Wang W., Nourshargh S., Voisin M.B. Эндогенный TNFα управляет переносом нейтрофилов в лимфатические сосуды и внутри них во время острого воспаления. науч. 2017;7:44189. [Бесплатная статья PMC] [PubMed] [Google Scholar]
науч. 2017;7:44189. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Баратин М., Форей С., Демария О., Хаббеддин М., Поллет Э., Маурицио Дж., Вертуй С., Давантуре С., Азукидзава Х., Флорес-Лангарика А. Гомеостатическая передача сигналов NF-κB в стационарных мигрирующих дендритных клетках регулирует иммунный гомеостаз и толерантность. Иммунитет. 2015;42:627–639. [PubMed] [Google Scholar]
Бертола А., Чиуччи Т., Руссо Д., Бурлье В., Дюффо К., Боннафус С., Блин-Ваккач К., Энти Р., Яннелли А., Гугенхайм Дж. Идентификация дендритных клеток жировой ткани коррелирует с резистентностью к инсулину, связанной с ожирением, и вызывает Th27-ответы у мышей и пациентов. Диабет. 2012;61:2238–2247. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Chawla A., Nguyen K.D., Goh Y.P. Воспаление, опосредованное макрофагами, при метаболических заболеваниях. Нац. Преподобный Иммунол. 2011; 11: 738–749.. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Чой Дж. Х., Бэнкс А. С. , Эстол Дж. Л., Каджимура С., Бострем П., Лазник Д., Руас Дж. Л., Чалмерс М. Дж., Каменецка Т. М., Блюхер М. Противодиабетические препараты ингибируют связанное с ожирением фосфорилирование PPARgamma с помощью Cdk5. Природа. 2010; 466: 451–456. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Эстол Дж. Л., Каджимура С., Бострем П., Лазник Д., Руас Дж. Л., Чалмерс М. Дж., Каменецка Т. М., Блюхер М. Противодиабетические препараты ингибируют связанное с ожирением фосфорилирование PPARgamma с помощью Cdk5. Природа. 2010; 466: 451–456. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Christodoulides C., Scarda A., Granzotto M., Milan G., Dalla Nora E., Keogh J., De Pergola G., Stirling H., Pannacciulli N ., Сетхи Дж.К. Мутации WNT10B при ожирении человека. Диабетология. 2006;49: 678–684. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Christodoulides C., Lagathu C., Sethi JK, Vidal-Puig A. Адипогенез и передача сигналов WNT. Тенденции Эндокринол. Метаб. 2009; 20:16–24. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Cipolletta D., Feuerer M., Li A., Kamei N., Lee J., Shoelson SE, Benoist C., Mathis D. PPAR-γ является основным драйвер накопления и фенотип клеток Treg жировой ткани. Природа. 2012; 486: 549–553. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Cipolletta D. , Cohen P., Spiegelman B.M., Benoist C., Mathis D. Появление и исчезновение сигнатуры мРНК, характерной для клеток Treg в висцеральной жировой ткани: возраст, диета и эффекты PPARγ. проц. Натл. акад. науч. США. 2015; 112: 482–487. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Cohen P., Spiegelman B.M., Benoist C., Mathis D. Появление и исчезновение сигнатуры мРНК, характерной для клеток Treg в висцеральной жировой ткани: возраст, диета и эффекты PPARγ. проц. Натл. акад. науч. США. 2015; 112: 482–487. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Corinti S., Albanesi C., la Sala A., Pastore S., Girolomoni G. Регуляторная активность аутокринного IL-10 в отношении функций дендритных клеток. Дж. Иммунол. 2001; 166:4312–4318. [PubMed] [Академия Google]
Cruz-Migoni S., Caamaño J. Ассоциированные с жиром лимфоидные скопления при воспалении и иммунитете. Фронт. Иммунол. 2016;7:612. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Damouche A., Lazure T., Avettan-Fènoël V., Huot N., Dejucq-Rainsford N., Satie A.P., Mélard A., David L., Gommet C., Ghosn J. Жировая ткань является запущенным вирусным резервуаром и местом воспаления при хронической ВИЧ- и SIV-инфекции. PLoS Патог. 2015;11:e1005153. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Де Смедт Т.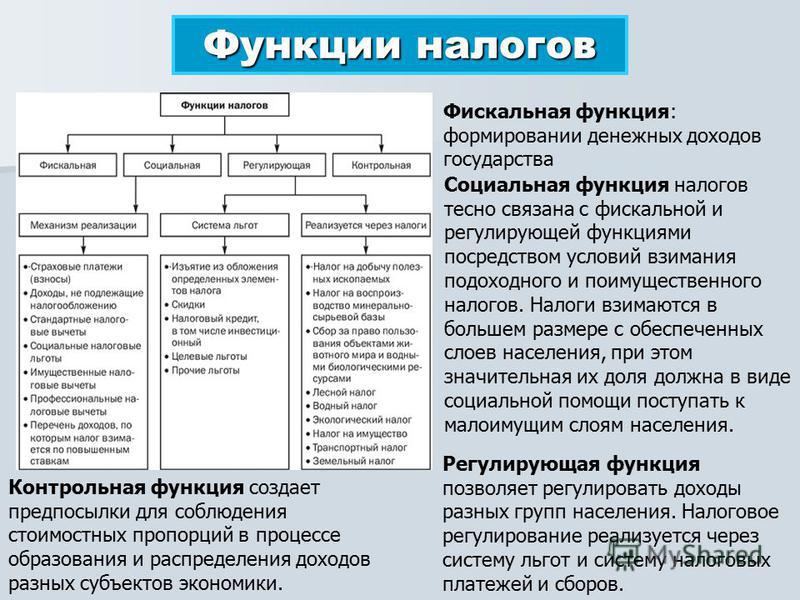 , Ван Мехелен М., Де Беккер Г., Урбен Дж., Лео О., Мозер М. Влияние интерлейкина-10 на созревание и функцию дендритных клеток. Евро. Дж. Иммунол. 1997; 27:1229–1235. [PubMed] [Google Scholar]
, Ван Мехелен М., Де Беккер Г., Урбен Дж., Лео О., Мозер М. Влияние интерлейкина-10 на созревание и функцию дендритных клеток. Евро. Дж. Иммунол. 1997; 27:1229–1235. [PubMed] [Google Scholar]
den Ruijter H.M., Pasterkamp G., de Jager S.C. Регуляция адипонектина при сердечно-сосудистых заболеваниях: показывает ли больной жир свой истинный цвет? Артериосклероз. тромб. Васк. биол. 2014;34:2180–2181. [PubMed] [Google Scholar]
Donath M.Y., Shoelson S.E. Сахарный диабет 2 типа как воспалительное заболевание. Нац. Преподобный Иммунол. 2011;11:98–107. [PubMed] [Google Scholar]
Донат М.Ю., Далмас Э., Заутер Н.С., Бони-Шнетцлер М. Воспаление при ожирении и диабете: дисфункция островков и терапевтические возможности. Клеточный метаб. 2013;17:860–872. [PubMed] [Google Scholar]
Farmer S.R. Транскрипционный контроль образования адипоцитов. Клеточный метаб. 2006; 4: 263–273. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Фонсека Д.М., Хэнд Т.В., Хан С.Дж. , Гернер М.Ю., Глатман Зарецкий А., Берд А.Л., Харрисон О.Дж., Ортис А.М., Хиноны М., Тринкьери Г. Зависит от микробиоты Последствия острой инфекции нарушают тканеспецифический иммунитет. Клетка. 2015; 163:354–366. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Гернер М.Ю., Глатман Зарецкий А., Берд А.Л., Харрисон О.Дж., Ортис А.М., Хиноны М., Тринкьери Г. Зависит от микробиоты Последствия острой инфекции нарушают тканеспецифический иммунитет. Клетка. 2015; 163:354–366. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Fu C., Liang X., Cui W., Ober-Blöbaum J.L., Vazzana J., Shrikant P.A., Lee K.P., Clausen B.E., Mellman I., Jiang A. β-катенин в дендритных клетках выполняет противоположные функции при перекрестном -примирование и поддержание Т-клеток CD8+ посредством регуляции ИЛ-10. проц. Натл. акад. науч. США. 2015;112:2823–2828. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Галаррага М., Кампьон Х., Муньос-Баррутиа А., Боке Н., Морено Х., Мартинес Х.А., Милагро Ф., Ортис-де-Солорсано К. Adiposoft: автоматизированное программное обеспечение для анализа клеточности белой жировой ткани в гистологических срезах. J. Липидный рез. 2012;53:2791–2796. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Гаскилл Б.Н., Гордон С. Дж., Пайор Э.А., Лукас Дж.Р., Дэвис Дж.К., Гарнер Дж.П. Тепло или изоляция: поведенческое титрование предпочтений мыши в отношении тепла или доступа к гнезду. ПЛОС Один. 2012;7:e32799. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Дж., Пайор Э.А., Лукас Дж.Р., Дэвис Дж.К., Гарнер Дж.П. Тепло или изоляция: поведенческое титрование предпочтений мыши в отношении тепла или доступа к гнезду. ПЛОС Один. 2012;7:e32799. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Gautier EL, Chow A., Spanbroek R., Marcelin G., Greter M., Jakubzick C., Bogunovic M., Leboeuf M., van Rooijen N., Хабенихт А.Дж. Системный анализ PPARγ в популяциях мышиных макрофагов выявил заметное разнообразие в экспрессии, играющее важную роль в разрешении воспаления и иммунитете дыхательных путей. Дж. Иммунол. 2012;189: 2614–2624. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Glass CK, Ogawa S. Комбинаторная роль ядерных рецепторов в воспалении и иммунитете. Нац. Преподобный Иммунол. 2006; 6: 44–55. [PubMed] [Google Scholar]
Grant S.F., Thorleifsson G., Reynisdottir I., Benediktsson R., Manolescu A., Sainz J., Helgason A., Stefansson H., Emilsson V., Helgadottir A. Вариант транскрипции Фактор 7-подобный ген 2 (TCF7L2) повышает риск развития диабета 2 типа.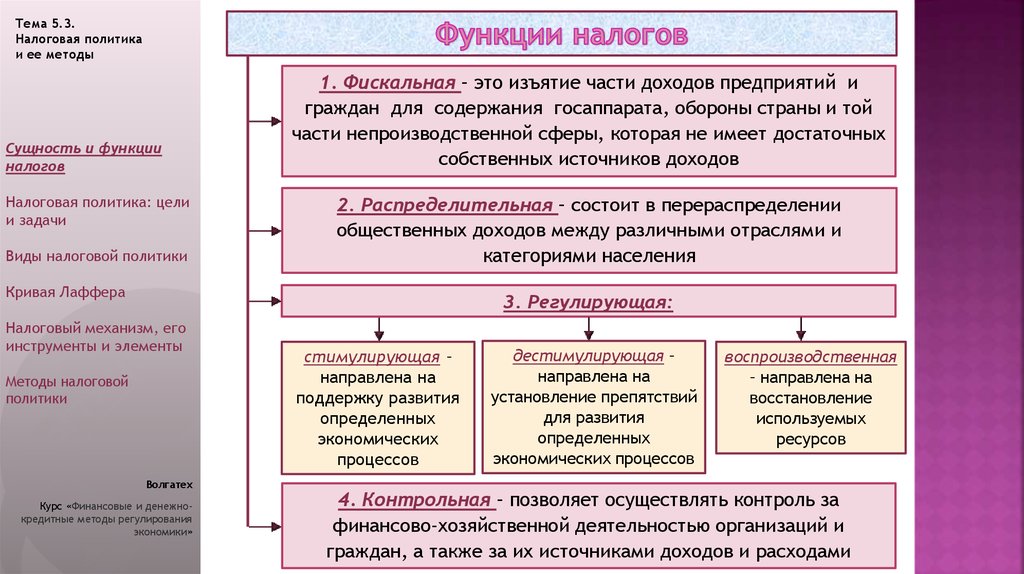 Нац. Жене. 2006; 38: 320–323. [PubMed] [Академия Google]
Нац. Жене. 2006; 38: 320–323. [PubMed] [Академия Google]
Iacobellis G., Bianco A.C. Эпикардиальная жировая ткань: возникающие физиологические, патофизиологические и клинические особенности. Тенденции Эндокринол. Метаб. 2011; 22: 450–457. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Клотц Л., Дани И., Эденхофер Ф., Нолден Л., Эверт Б., Пол Б., Коланус В., Клокгетер Т., Кнолле П., Диль Л. Гамма-контроль рецептора, активируемого пролифератором пероксисом, функции дендритных клеток способствует развитию анергии CD4+ Т-клеток. Дж. Иммунол. 2007;178:2122–2131. [PubMed] [Академия Google]
Комия Ю., Хабас Р. Пути передачи сигнала Wnt. Органогенез. 2008; 4: 68–75. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Куан Э.Л., Иванов С., Бриденбо Э.А., Виктора Г., Ван В., Чайлдс Э.В., Платт А.М., Якубзик С.В., Мейсон Р.Дж., Гашев А.А. Повышение проницаемости лимфатических сосудов способствует воспалению жировой ткани и распространению антигена в дендритные клетки жировой ткани, направляющие лимфатические узлы. Дж. Иммунол. 2015;194:5200–5210. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Дж. Иммунол. 2015;194:5200–5210. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Лю Дж., Ван Х., Цзо Ю., Фармер С.Р. Функциональное взаимодействие между гамма-рецептором, активируемым пролифератором пероксисом, и бета-катенином. Мол. Клетка. биол. 2006; 26: 5827–5837. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Лошко Дж., Шрайбер Х.А., Рике Г.Дж., Эстерхази Д., Мередит М.М., Педикорд В.А., Яо К.Х., Кабальеро С., Памер Э.Г., Муцида Д., Нуссенцвейг М.К. Отсутствие МНС класса II на кДК приводит к микробно-зависимому воспалению кишечника. Дж. Эксп. Мед. 2016; 213:517–534. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Manicassamy S., Reizis B., Ravindran R., Nakaya H., Salazar-Gonzalez R.M., Wang YC, Pulendran B. Активация бета-катенина в дендритных клетках регулирует иммунитет по сравнению с толерантностью в кишечнике. Наука. 2010; 329:849–853. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Manoharan I., Hong Y., Suryawanshi A., Angus-Hill ML, Sun Z.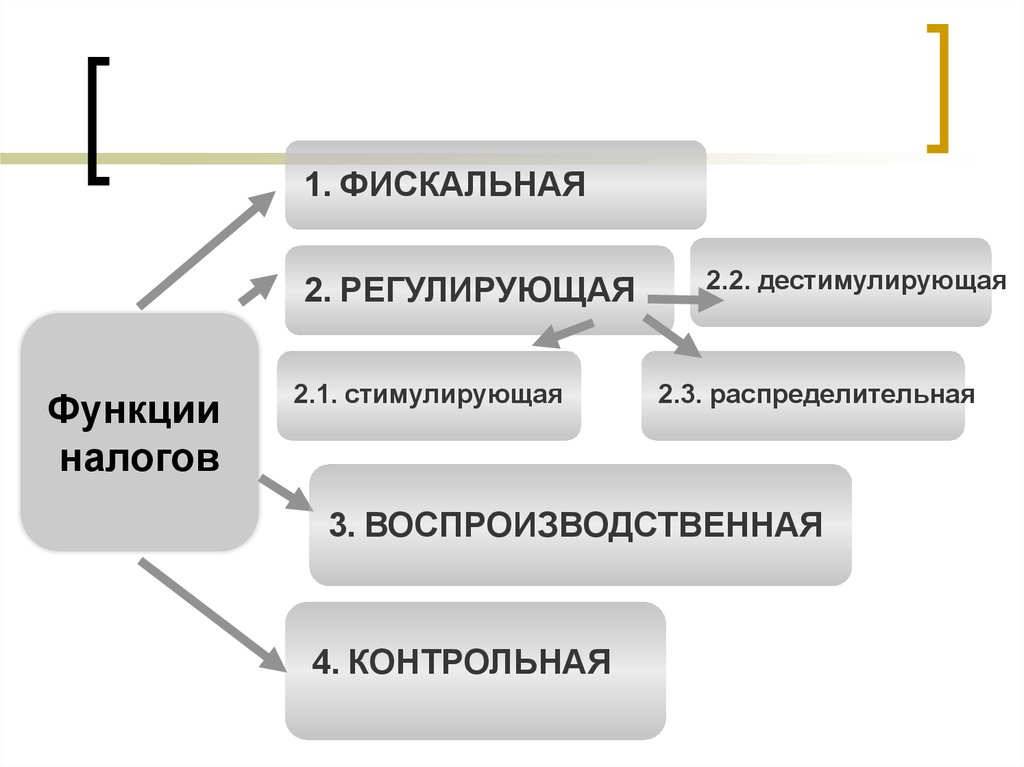 , Mellor AL, Munn D.H., Manicassamy S. TLR2-зависимая активация β -катениновый путь в дендритных клетках индуцирует регуляторные ответы и ослабляет аутоиммунное воспаление. Дж. Иммунол. 2014;193:4203–4213. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Mellor AL, Munn D.H., Manicassamy S. TLR2-зависимая активация β -катениновый путь в дендритных клетках индуцирует регуляторные ответы и ослабляет аутоиммунное воспаление. Дж. Иммунол. 2014;193:4203–4213. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Mattacks C.A., Sadler D., Pond C.M. Влияние пищевых липидов на дендритные клетки перинодальной жировой ткани при хроническом легком воспалении. бр. Дж. Нутр. 2004; 91: 883–892. [PubMed] [Google Scholar]
Menezes S., Melandri D., Anselmi G., Perchet T., Loschko J., Dubrot J., Patel R., Gautier E.L., Hugues S., Longhi M.P. Гетерогенность моноцитов Ly6Chi контролирует их дифференцировку в макрофаги iNOS+ или дендритные клетки, происходящие из моноцитов. Иммунитет. 2016;45:1205–1218. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Мерад М., Сате П., Хелфт Дж., Миллер Дж., Морта А. Линия дендритных клеток: онтогенез и функция дендритных клеток и их подмножеств в стабильном состоянии и в условиях воспаления. Анну. Преподобный Иммунол. 2013; 31: 563–604. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Преподобный Иммунол. 2013; 31: 563–604. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Nguyen MT, Chen A., Lu WJ, Fan W., Li PP, Oh D.Y., Patsouris D. Регуляция экспрессии хемокинов и хемокиновых рецепторов с помощью PPARγ в адипоцитах и макрофагах . ПЛОС Один. 2012;7:e34976. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Odegaard J.I., Ricardo-Gonzalez R.R., Goforth M.H., Morel C.R., Subramanian V., Mukundan L., Red Eagle A., Vats D., Brombacher F., Ferrante A.W., Chawla A. Макрофаг-специфический PPARgamma контролирует альтернативную активацию и улучшает резистентность к инсулину. Природа. 2007; 447:1116–1120. [PMC free article] [PubMed] [Google Scholar]
Окин Д., Меджитов Р. Влияние длительного воспаления на печеночный мевалонатный путь приводит к гипергликемии. Клетка. 2016; 165:343–356. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Паскуаль Г., Фонг А.Л., Огава С., Гамлиэль А., Ли А.К., Перисси В., Роуз Д.В., Уилсон Т.М., Розенфельд М. Г., Гласс К.К. Зависимый от SUMOylation путь опосредует трансрепрессию генов воспалительного ответа с помощью PPAR-гамма. Природа. 2005; 437: 759–763. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Г., Гласс К.К. Зависимый от SUMOylation путь опосредует трансрепрессию генов воспалительного ответа с помощью PPAR-гамма. Природа. 2005; 437: 759–763. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Пацурис Д., Ли П.П., Тапар Д., Чепмен Дж., Олефски Дж.М., Нильс Дж.Г. Абляция CD11c-позитивных клеток нормализует чувствительность к инсулину у тучных животных с резистентностью к инсулину. Клеточный метаб. 2008; 8: 301–309.. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Pond C.M., Mattacks C.A. Источник жирных кислот, встроенных в пролиферирующие лимфоидные клетки в иммуностимулируемых лимфатических узлах. бр. Дж. Нутр. 2003; 89: 375–383. [PubMed] [Google Scholar]
R Основная команда. (2016). R: язык и среда для статистических вычислений (Вена, Австрия).
Рикот М., Ли А.С., Уилсон Т.М., Келли С.Дж., Гласс С.К. Гамма-рецептор, активируемый пролифератором пероксисом, является негативным регулятором активации макрофагов. Природа. 1998;391:79–82. [PubMed] [Google Scholar]
Ричи М. Е., Фипсон Б., Ву Д., Ху Ю., Лоу К.В., Ши В., Смит Г.К. limma обеспечивает анализ дифференциальной экспрессии для секвенирования РНК и исследований микрочипов. Нуклеиновые Кислоты Res. 2015;43:e47. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Е., Фипсон Б., Ву Д., Ху Ю., Лоу К.В., Ши В., Смит Г.К. limma обеспечивает анализ дифференциальной экспрессии для секвенирования РНК и исследований микрочипов. Нуклеиновые Кислоты Res. 2015;43:e47. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Росс С.Э., Хемати Н., Лонго К.А., Беннетт С.Н., Лукас П.С., Эриксон Р.Л., Макдугалд О.А. Ингибирование адипогенеза посредством передачи сигналов Wnt. Наука. 2000; 289:950–953. [PubMed] [Google Scholar]
Шиппер Х.С., Праккен Б., Калховен Э., Боэс М. Иммунные клетки, резидентные в жировой ткани: ключевые игроки в иммунометаболизме. Тенденции Эндокринол. Метаб. 2012; 23:407–415. [PubMed] [Академия Google]
Шрайбер Х.А., Лошко Дж., Карссемейер Р.А., Эсколано А., Мередит М.М., Муцида Д., Гермонпрез П., Нуссенцвейг М.К. Кишечные моноциты и макрофаги необходимы для поляризации Т-клеток в ответ на Citrobacter rodentium. Дж. Эксп. Мед. 2013;210:2025–2039. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Silva-García O. , Valdez-Alarcón J.J., Baizabal-Aguirre V.M. Сигнальный путь Wnt/β-катенин контролирует воспалительную реакцию при инфекциях, вызванных патогенными бактериями. Медиаторы воспаления. 2014;2014:310183. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Valdez-Alarcón J.J., Baizabal-Aguirre V.M. Сигнальный путь Wnt/β-катенин контролирует воспалительную реакцию при инфекциях, вызванных патогенными бактериями. Медиаторы воспаления. 2014;2014:310183. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Стефанович-Рачич М., Ян X., Тернер М.С., Мантелл Б.С., Штольц Д.Б., Самптер Т.Л., Сипула И.Дж., Дедузис Н., Скотт Д.К., Морел П.А. Дендритные клетки способствуют инфильтрации макрофагов и составляют значительную часть связанного с ожирением увеличения количества клеток CD11c+ в жировой ткани и печени. Диабет. 2012;61:2330–2339. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Suryawanshi A., Manoharan I., Hong Y., Swafford D., Majumdar T., Taketo M.M., Manicassamy B., Koni P.A., Thangaraju M., Sun Z Каноническая передача сигналов wnt в дендритных клетках регулирует ответы Th2/Th27 и подавляет аутоиммунное нейровоспаление. Дж. Иммунол. 2015;194:3295–3304. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Тан В., Зеве Д. , Су Дж. М., Боснаковски Д., Киба М., Хаммер Р. Э., Таллквист М. Д., Графф Дж. М. Клетки-предшественники белого жира находятся в жировой сосудистой сети. . Наука. 2008; 322: 583–586. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Су Дж. М., Боснаковски Д., Киба М., Хаммер Р. Э., Таллквист М. Д., Графф Дж. М. Клетки-предшественники белого жира находятся в жировой сосудистой сети. . Наука. 2008; 322: 583–586. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Тановиц Х.Б., Шерер П.Е., Мота М.М., Фигейредо Л.М. Жировая ткань: убежище для паразитов? Тенденции Паразитол. 2017; 33: 276–284. [Статья бесплатно PMC] [PubMed] [Google Scholar]
Wang F., Mullican S.E., DiSpirito J.R., Peed LC, Lazar M.A. Липоатрофия и тяжелые метаболические нарушения у мышей с жироспецифической делецией PPARγ Proc. Натл. акад. науч. США. 2013;110:18656–18661. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Ван К.А., Тао С., Гупта Р.К., Шерер П.Е. Отслеживание адипогенеза при развитии, увеличении и регенерации белой жировой ткани. Нац. Мед. 2013;19:1338–1344. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Weber C., Meiler S., Döring Y., Koch M., Drechsler M., Megens R.T., Rowinska Z., Bidzhekov K., Fecher C.